- +1
基因编辑,魔剪还是魔鬼?| 诺奖背后
Jennifer Doudna和Emmanuelle Charpentier获得2020年诺贝尔化学奖,被很多人认为实至名归,张锋再次错过大奖,虽然有争议,但是,总的来说,理论创新是第一位的,行动是第二位的。这篇文章综合了多篇网络文章,以及增加了一些最新的研究,概括了基因编辑的系统发现到2020年的主要的里程碑。
重大的科学发现经常是站在巨人肩上获得的,在CRISPR基因编辑技术发现史上,还有很多英雄,虽然他们没有获得诺奖,但是他们的贡献是伟大的。关于CRISPR基因编辑技术发现史有大量的综述或文章,其中最有影响的还是2016年1月14日,Cell杂志发表的美国哈佛-MIT Broad研究所教授Eric Lander撰写的一篇关于CRISPR基因编辑技术发现史的综述文章,虽然,这篇文章发表后,受到Jennifer Doudna公开的驳斥,但是不容否认这篇文章揭密了基因编辑道路中的英雄故事。
生物学家们一直在打磨能够进行DNA编辑的工具,TALEN,ZFN都只是过客,而CRISPR技术很快成为了其中最耀眼的明星。CRISPR体系包括一个细菌核酸酶(Cas)和一段与目标DNA匹配的引导RNA,能为细菌沉默入侵者 (比如病毒)遗传信息的关键部分。与其他基因编辑技术相比,CRISPR技术更易于操作扩展性也更强,因此迅速成为了科研领域的宠儿,为基因工程和生物医学领域带来了一场革命。
很少有发现能够像CRISPR那样在一夜之间改变整个领域。科学家们三年前才发现CRISPR能在活细胞中实现精确有效的基因组编辑。CRISPR/Cas免疫系统主要有三种类型,而这项研究使用的主要是完全依赖Cas9内切酶进行靶标和 剪切的II型CRISPR系统。自此以后,CRISPR技术就如风暴一般席卷了整个科研领域,现在正有数以千计的实验室在使用它。
CRISPR技术的应用范围非常广泛,比如建立人类遗传病和癌症的复杂动物模型、进行人类细胞全基因组筛选以鉴定生物学过程背后的基因、启动或关闭特定的基因,设计转基因植物等等。CRISPR改造人类生殖细胞的可能性更是引起了广泛的争议。
想必现在已经没有哪个分子生物学家还未听过CRISPR的大名。然而大部分人可能还不清楚这一革命到底是如何发生的。与过程相比,做科研的人似乎更注重结果。一旦某个事实被牢固地建立起来,背后的曲折路径就显得不那么重要了。
Cell亮点文章:CRISPR英雄谱
著名遗传学家、美国科学院院士Eric S. Lander教授(麻省理工学院与哈佛大学Broad研究所主席兼主任)却不这么看。他认为,了解科研进展背后的人和事能让我们获益良多。对于迈入科研门槛不久的学生来说,对科学发现有一个真实的概念特别重要,不仅有重要的指导意义还能带来关键性的启发。

CRISPR的发现者:Francisco Mojica CRISPR
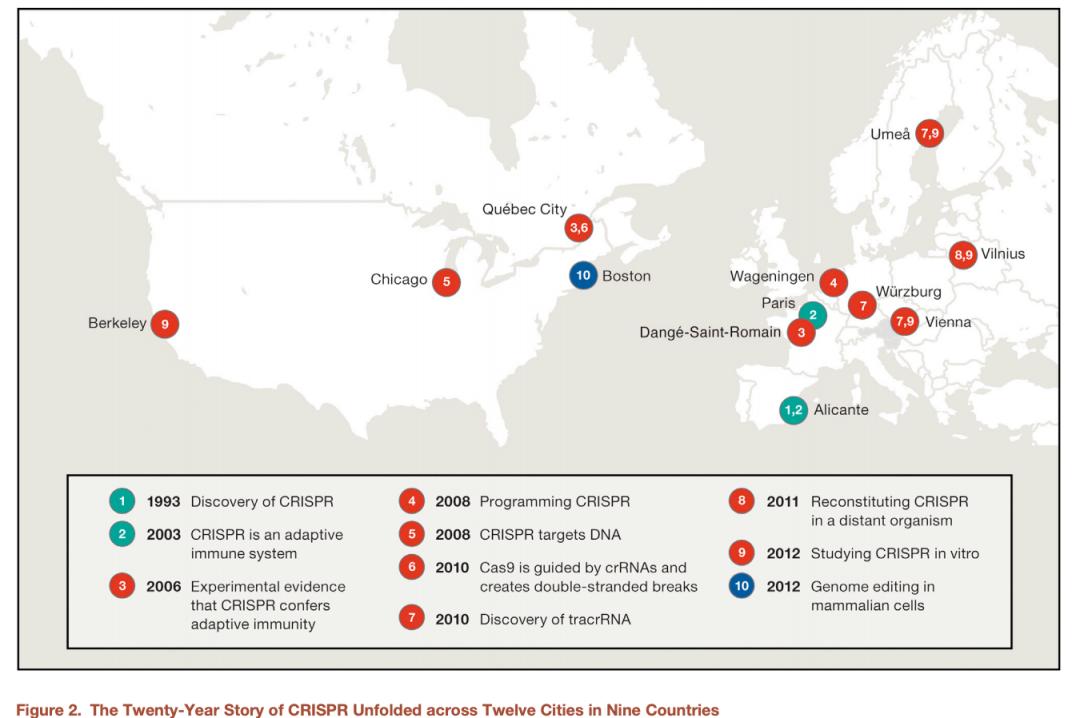
这位28岁的博士研究生被这种神秘的结构迷住了,在此后十年的职业生涯中一直致力于揭开这一谜团。Mojica很快发现类似的重复也存在于其他一些不同种类的细菌中。他意识到这样的结构可能在原核生物中具有重要的功能。他发表了一篇论文,报告了这类新的重复序列(Mojica et al., 1995)。
当Mojica在阿利坎特大学任职的时候,由于缺少启动资金和实验室空间,他主要通过生物信息学手段来研究这种奇怪的重复。Mojica最初将其命名为SRSR(short regularly spaced repeats),后来又将名字改为CRISPR(规律成簇的间隔短回文重复)(Jansen 等, 2002; Mojica 和 Garrett, 2012)。
到2000年,Mojica已在20个不同的微生物上发现了CRISPR基因位点,其中包括结核分枝杆菌(Mycobacterium tuberculosis),艰难梭菌(Clostridium difficile),和鼠疫杆菌(Yersinia pestis)(Mojica 等, 2000).。在两年的时间内,研究人员已经将这相关微生物的名录翻倍并且列录了基因位点的关键特征,包括存在近亲缘的特定的“CRISPR关联基因”(cas基因),这通常认为与他们的功能相关(Jansen 等, 2002)。(表1列举了CRISPR系统的最新分类。)


但是,CRISPR系统的功能究竟是什么呢?各种假设层出不穷:比如设想与基因调控,复制分区,DNA修复还有其它功能有关。然而大多数的这类假设都没有证据支持,它们一个一个都被证伪了。如同CRISPR的发现一样,重要的睿见来自于生物信息学。
CRISPR是一种适应性免疫系统
2003年8月的假期,Mojica 避开圣波拉的酷暑,躲在阿利坎特的空调办公室里。如今已然作为初兴的CRISPR领域的领军人物,他将目光从重复序列转向分隔它们的间隔序列。使用文字处理器,Mojica 不辞辛劳地抽出每一个间隔并将其插入BLAST软件来搜索与其它任何已知DNA序列的相似性。
虽然他尝试这种方法失败过,但是DNA序列数据库在不断扩大,这一次他成功掘到金矿了。在他最近从一种大肠杆菌菌株测序到的一个CRISPR基因位点上,其中一个间隔与一种P1噬菌体的序列相匹配,而这一噬菌体可以感染多种大肠杆菌菌株。然而,携带这一间隔的菌株已知对P1感染具有抵抗力。那一周结束时,他已经检索了4500个间隔。其中88个间隔与已知序列相似,三分之二同携带间隔的微生物相关的病毒或接合质粒相匹配。Mojica意识到CRISPR基因位点储存了用于为保护微生物抵抗感染的适应性免疫系统所需的信息。
Mojica 于是和同事们一起外出饮干邑白兰地庆祝,并在第二天清晨开始撰写相关论文。如此竟开始了长达18个月的痛苦煎熬。认识到这一发现的重要性,Mojica 将论文投给了《自然》(Nature)。2003年11月期刊在未征询外部评审的情况下拒绝的论文的发表。难以理解的是,编辑声称论文的关键论点属于已知范畴。2004年1月,《美国国家科学院院刊》(PNAS)的决定是这篇论文缺乏“充足的新颖观点和重要性”因而不够资格送审。《分子微生物学》和《核酸研究》(NAR)也相继地拒绝发表。此时,绝望而又担心被别人抢先一步发表的Mojica将论文投给了《分子演化杂志》。在经过12个多月的审稿和修订,这篇宣布CRISPR可能功能的论文终于在2005年2月1日发表了(Mojica et 等, 2005)。

与此同时,CRISPR正是另一个意想不到的地点的研究人员的关注焦点,那是巴黎向南30公里以外,法国国防部的一个研究部门。Giles Vergnaud 是在巴斯德研究所受训的人类遗传学家,他的博士和博士后研究都受到法国武器装备总局的资助。他在1987年完成博士后研究之后加入法国国防部并建立它的第一个分子生物学实验室。
在以后的十年间,Vergnaud继续着人类遗传学的研究工作。但是,情报部门在九十年代后期关于伊拉克的萨达姆·萨达姆政权正在发展生物武器的报告引起关切之后,国防部在1997年要求Vergaud和他的团队将研究重心转向法医微生物学,籍此发展基于微生物种类间细微遗传学差别来追踪病原体来源的手段。在与附近的巴黎第十一大学的遗传与微生物学研究所的联合实验室,他开始使用串联重复序列多态性,这一法医学人类DNA指纹图谱的主要工具,来绘制导致炭疽病(anthrax)和鼠疫的细菌种类。
法国国防部拥有一批特殊的来自1964至1965年越南鼠疫爆发期间的61件鼠疫杆菌样本。Vergnaud发现这些密切相关联的分离株在串联重复基因位点上是一致的,除去一个位置是例外,即由他的同事Christine Pourcel所发现的CRISPR基因位点。它们的品系由偶尔出现新间隔所区分,而这些间隔无不是在CRISPR基因位点的前段末端(Pourcel 等,2005)。
值得注意的是,其中许多的新间隔与存在于鼠疫杆菌基因上的原噬菌体(prophage)相配。作者们推断,CRISPR基因位点是在执行防御机制,用诗化的语言来说,就是“CRISPR可能重现‘过往遗传攻击’的记忆。”Vergnaud试图发表他们发现的努力和Mojica遇到了相同的阻碍。论文被《美国国家科学院院刊》、《细菌学学刊》、《核酸研究》以及《基因组研究》相继退稿,直到在2005年3月1日的《微生物学》上发表。


John van der Oost于1989年在阿姆斯特丹自由大学(Free University of Amsterdam)获得博士学位,他最初志在解决世界清洁能源的需求而研究如何利用蓝藻来生产生物燃料。在返回阿姆斯特丹之前,他相继在赫尔辛基和海德堡研究细菌的代谢途径。1995年,瓦赫宁根大学(Wageningen University)向他发出终身教职的聘任邀请,但是条件是需要由他来壮大一个专攻极端条件生存的微生物的团队。
Van der Oost 在德国期间已经听说过能够在黄石国家公园的温泉中繁衍的嗜热链球菌,于是对探索这些奇特微生物在代谢途径上的演化区别跃跃欲试。他开始和隶属于美国国家卫生研究所(National Institutes of Health)的国家生物科技信息中心(NCBI)的微生物进化和计算机生物学领域的专家Eugene Koonin合作。Koonin早已开始对CRISPR系统进行数据分类和分析,他在2005年的一次访问中将van der Oost领入当时还鲜为人知的CRISPR领域(Makarova等,2006)。
Van der Oost当时已从荷兰国家科学基金获得了主要资助。他决定在研究申请书的课题之外,将部分经费用作CRISPR的研究。
他和同事们将一种大肠杆菌的CRISPR系统嵌入另一缺乏这一自身内源性系统的大肠杆菌品系。这样就可以使他们可以在生物化学上鉴定一组拥有5个cas蛋白的复合体,它被称作Cascade(Brouns等,2008)。(大肠杆菌拥有更复杂的1类,I 型CRISPR系统,其中cas9的功能是由Cascade复合体联同核酸酶cas3一起来实现的。见表1。)
通过将每一个组成部分逐一剔除,他们证明Cascade是将经由CRISPR基因位点转录的一个长前体RNA接入61个核苷酸长的CRISPR RNA(crRNAs)所必需的。经过对一组与Cascade复合体一同纯化的crRNAs进行克隆和基因测序,发现它们都从8个碱基的重复序列开始,紧随其后的是完全间隔和新重复区的出现。这一发现支持了先前的假设,即重复序列的回文(palindromic)结构特性导致crRNA中次级结构的形成(Sorek等,2008)。
为了证明crRNA序列是产生基于CRISPR的抵抗力的原因,他们着手创造首个人工CRISPR排列,设定使CRISPR将λ噬菌体的四种基本基因作为定向靶位。正如他们所料,携带新的CRISPR序列的品系对噬菌体呈抵抗性。这是有史以来第一个直接由程序设计的基于CRISPR的免疫,对细菌而言犹如流感疫苗。
这些实验结果暗示,CRISPR的目标不是RNA(Bolotin所设想的),而是DNA。研究人员设计了两种版本的CRISPR排列,一种是反义链方向的(与mRNA和DNA位点的编码链互补),另一种是正义链方向的(仅仅与另一条DNA链互补)。虽然,间隔在有效性上各有差别,但是实验在正义链方向的版本上起了效果这一事实有力说明目标不是mRNA。但是,它不是直接证据。在《科学》的编辑们针对论文作出坚定论断保持谨慎的要求下,van der Oost将CRISPR定位DNA为目标物的观点以“猜想”的方式提出。

Luciano Marraffini正在芝加哥大学完成博士学位的研究,研究方向是葡萄球菌(Staphylococcus),此时从系里的噬菌体遗传学世界权威Malcolm Casadaban那里了解到CRISPR。Casadaban于2005年旋即看出CRISPR有可能是适应性免疫系统这一发现的重要程度,而且对所有对此表现出兴趣的人都谈论CRISPR。和许多噬菌体研究领域的科学家一样,Marraffini坚信CRISPR不是由RNA干扰来作用的,因为这一机制对于克服发生在噬菌体感染过程中的爆发式增长是无能为力的。他推断,CRISPR必然对DNA进行了切割,这个功能就好像限制性内切酶的作用。
Marraffini本来热切希望加入世界上为数不多的几个正在研究CRISPR的研究团队来做博士后的研究,可是由于他的妻子在库克县的伊利诺伊(Cook County, Illinois)刑事法庭有一个做翻译员的好工作,他感觉自己必须留在芝加哥。他说服西北大学(Northwestern University)的生物化学家Erik Sontheimer让自己加入他的实验室研究CRISPR,Sontheimer一直从事RNA剪接和RNA干扰的工作。
早在搬去西北大学之前,Marraffini在完成博士研究同时就开始了关于CRISPR的工作,研究葡萄球菌(Staphylococcus)的CRISPR系统是否可以阻止质粒接合。他注意到,一种表皮葡萄球菌(Staphylococcus epidermidis)拥有这样一个间隔,它与来自具有抗生素耐药性的金黄色葡萄球菌的质粒上存储的切口酶(nickase,nes)基因的某区域相匹配。他证明了,这些质粒不能转化表皮葡萄球菌,而中断质粒内nes序列或者它在CRISPR基因位点上的相配间隔的两者之一都会取消干扰功能(Marraffini 和 Sontheimer,2008)。显而易见,CRISPR如同阻止病毒一样阻止了质粒。
Marraffini和Sontheimer一度考虑在试管里重组CRISPR系统来证明它可以切割DNA。但是表皮葡萄球菌的系统太过复杂,它有9个cas基因,并且它的基因特点还完全未被掌握。于是,他们将目光转向分子生物学。他们聪明地对CRISPR系统定向射靶的质粒中的nes基因进行了改变,就是通过在它的序列中间嵌入一个自我剪接的内含子。如果CRISPR是以mRNA为目标的话,那么这一变化就不会影响干扰功能,因为内含子序列会被剪除。而如果CRISPR是以DNA为目标的话,那么这种嵌入就会取消干扰功能因为间隔将不再匹配。结果已经清楚:CRISPR的目标是DNA。
Marraffini和Sontheimer认识到CRISPR在本质上是一种可编辑设定的限制性内切酶。他们的论文则第一个清楚提出了预测:CRISPR可以被转而用于在异源性系统内进行基因组编辑。“从实践的观点来看,” 他们宣布,“可以对包含任何已知24至48个核苷酸目标物的DNA进行可定位灭除的导向能力具有可观的功能性用途,特别是如果这个系统还可以在原来的细菌或古菌环境以外发挥作用。”他们甚至提交了涉及使用CRISPR在真核细胞内切割和纠正基因位点的专利申请,然而由于尚缺乏实验证明而最终放弃了申请(Sontheimer和Marraffini,2008)。
Cas9是由crRNAs指引并且在DNA中产生双链裂断
在2007年影响深远的研究确认了CRISPR是一种适应性免疫系统之后(Barrangou等,2007),Sylvain Moineau继续与丹尼斯科公司合作来弄清CRISPR切割DNA的机制。
问题是CRISPR在通常情况下总是如此高效,使得Moineau和同事们没法轻易观察到入侵的DNA是如何被消灭的。但在研究嗜热链球菌的质粒干扰过程中他们受到了幸运女神的眷顾。研究人员发现了少量品系,它们的CRISPR对依靠电穿术(electroporation)转化的质粒仅仅可以提供部分防卫。在其中一个低效的品系的细胞内,可以看见呈线性的质粒依旧存在。一定程度上,质粒干扰的过程能够被减缓到使CRISPR活动的产物可以被观察到(Garneau等,2010)。
这一品系使他们得以研究切割的过程。与他们先前的结果一致(Barrangou等,2007),这次的结果显示质粒的切割依赖cas9核酸酶。当对线性化的质粒进行测序时,他们在PAM(proto-spacer adjacent motif)的上游发现了单个精准平端切割的三核苷酸,PAM是一种关键性序列特征,而先前的论文对它的作用已有描述(Deveau等,2008;Hovath等,2008)。将分析的范围扩展之后,结果表明病毒的DNA也是在与PAM序列有关的相同位置被精准切割的。此外,匹配同一目标的不同间隔的数量与切口数量一致。
实验结果确定无疑地表明Cas9的核酸酶活性是在精准的点位上对DNA进行切割的,而这些点位则由crRNA的特定序列编码。
tracrRNA的发现
即使在对CRISPR-Cas9系统的密集研究之下,组成这个难题一块拼图却仍然缺失,就是后来被称作反式作用CRISPR RNA(tracrRNA)的一个小RNA。事实上,它的发现者Emmanuelle Charpentier(诺奖得主之一)和Jörg Vogel并不是专门研究CRISPR系统的;他们仅仅试图鉴定微生物RNA。
Charpentier于1995年从巴斯德研究所(Pasteur Institute)获得博士学位,之后在纽约做了6年的博士后研究,然后相继于2002年在维也纳大学和2008年在瑞典的于默奥大学(UmeåUniversity)建立自己的实验室。她在化脓性链球菌(Streptococcus pyogenes)里发现了一种不寻常的可控制其传染力的RNA之后(Mangold等,2004)开始对鉴定更多微生物内的调控性RNA发生了兴趣。她用生物信息学的程序检索化脓性链球菌的基因区间,设想它们可能编码非编码RNA。她发现了若干候选区间,包括在CRISPR基因位点附近的一个,——但是缺乏关于这些RNA的直接信息,就太难继续研究了。
当Charpentier在威斯康辛的麦迪逊(Madison, Wisconsin)召开的2007年RNA学会上遇见Vogel时,解决方法出现了。在德国受训为微生物学家的Vogel在乌普萨拉(Uppsala)和耶路撒冷(Jerusalem)做博士后研究期间开始专攻在病原体内寻找RNA,这一工作持续至2004年他在位于柏林的马普感染生物学研究所建立自己的研究小组时。(5年之后,他转去乌兹堡(Würzburg)领导一个传染病研究中心。)随着“下一代测序”技术的到来,Vogel意识到大规模平行测序将使得绘制任何微生物转录组的整体图录成为可能。他当时已经将这种方法用于导致胃溃疡的细菌——幽门螺杆菌(Sharma等,2010),同时正在用于其它细菌。Charpentier和Vogel决定将目标瞄准化脓性链球菌。
此方法产生了一个惊人的结果:仅次于核糖体RNA和转运RNA之后数量第三多的RNA转录本属于一类新的小RNA,它是从紧挨着CRISPR基因位点(就是那个曾引起Chapentier注意的区间)的序列转录过来的,并且它有25个与CRISPR重复序列近乎完美地互补的碱基。这一互补性说明tracrRNA和crRNA的前体一起杂交并且被RNaseIII切割处理成为成熟产物。遗传删除实验也证实了这个观点,即tracerRNA对处理crRNA是必不可少的,因而就对CRISPR作用也是必不可少(Deltcheva 等, 2011)。

在远缘物种内重建CRISPR
Virginijus Siksnys成长于苏联时代的立陶宛(Lithuania),从维尔纽斯大学(Vilnius University)毕业之后于1980年代早期前往莫斯科国立大学(Moscow State University)攻读博士学位,研究酶动力学。这之后他回到家乡维尔纽斯加入应用酶学研究所,从事当时热门的限制性内切酶领域的研究。20年之后,他对于研究限制性内切酶已经感到厌烦。Horath,Barrangou和Moineau的2007年论文重燃了他对于细菌抵御外部DNA的屏障的兴趣。作为化学家,他意识到想要理解CRISPR,就必须在体外重建它。
他的第一步就是要测试他是否已经获取了所有的必要组成部分。他和合作者们着手观察来自嗜热链球菌(S. thermophiles)的CRISPR基因位点是否能够在一个远缘的微生物大肠杆菌(E. coli)内被重建成为完全发挥作用的形态。令他们感到欣喜的是,他们发现转入整个CRISPR基因位点就足以实现对质粒和噬菌体DNA的靶向干扰(Sapranauskas 等,2011)。他们用异源系统还证明了Cas9是干扰活动唯一必需的蛋白,而它的Ruvc和HNH核酸酶结构域都是必不可少的。
随着CRISPR-Cas9干扰系统的必要与充分的组成部分——Cas9 核酸酶,crRNA和tracrRNA都被发现了,这一领域的研究到达了一个关键性的里程碑。这个系统依靠尖端的生物信息学、遗传学和分子生物学已经被化整为零地完全掌握了。如今是时候将方向转向精确的生物化学实验来证实并且在试管里拓展这些结论了。
在试管里研究CRISPR
利用它们在大肠杆菌里的异源表达系统,Siksnys和同事们通过使用链酶亲和素标记物标记Cas9来纯化嗜热链球菌的Cas9-crRNA复合体,并在在试管内观察它的活动(Gasiunas 等,2012)。结果表明,复合体能够在体外切割DNA目标,创造出一个距离PAM序列恰好3个核苷酸的双链断裂——正好与Moineau及其同事在细菌体内观察到的现象一致。最重要的是,实验证明他们能够在CRISPR阵列里对Cas9和专门设计的间隔序列一同改编,从而实现在试管内对选择目标位置的切割。
通过转变HNH与RuvC核酸酶结构域的催化残基,他们还证明了前者切割与crRNA互补的基因链,而后者则切割与之相对的那条链。他们同时又证明了crRNA可以被修剪至只剩20个核苷酸而仍然保持有效切割力。最后,Siksnys证明这个系统还可以用第二种方法重建——就是把纯化的His标记的Cas9、在试管内转录的tracrRNA和crRNA,还有RNase III混合起来——而上述两种RNA对于Cas9切割DNA都是必不可少。(最终他们在修订后的论文里删除了第二种重建方法,但是在他们已发表的提交于2012年3月的美国专利申请书中报告了所有的研究内容[Siksnys 等,2012])。

夏威夷长大的Doudna在哈佛大学获得博士学位,与Jack Szostak合作将一个RNA自我剪接内含子重塑为具有复制RNA模版能力的核酶。她在之后跟科罗拉多大学的Tom Cech做博士后研究时解决了核酶的晶体结构问题。在她自己的实验室(起先于1994年在耶鲁建立,之后于2002年在伯克利),她鉴定了多种现象下的RNA蛋白复合体,诸如与内部核糖体入口位置有关和microRNA的加工有关等。她一直使用晶体学和冷冻电镜来解决I型CRISPR系统的Cascade复合体的组成部分的结构问题,这是一种用于诸如大肠杆菌的较为复杂的系统。
两位科学家于是决定联手。她们使用重组Cas9(来自在大肠杆菌里表达的化脓性链球菌[S. pyogenes]的基因)和体外转录的crRNA及tracrRNA(Jinek 等,2012)。和Siksnys一样,她们也证明了Cas9可以在体外切割纯化的DNA,它可以和专门设计的crRNA一起被设计,两种核酸酶结构域分别切割相对的两条DNA链,并且crRNA和tracrRNA对Cas9发挥作用都是必需的。此外,她们证明两种RNA在被融合为单一的导向RNA(sgRNA)时也可以在体外发挥作用。在经过其他科学家的修改而变得可以更高效地在体外作用之后,sgRNA的概念在基因组编辑领域被广泛使用。


在哺乳动物细胞内进行基因组编辑
在1980年代后期科学家们设计出一种可以在活细胞内改变哺乳动物基因的方法,这彻底改变了生物医学研究,包括使得在老鼠的胚胎干细胞内的特定位置嵌入DNA并且培育出携带这一遗传改变的后代成为可能(综述见Capeccchi,2005)。虽然这个方法是革命性的,但是过程却是低效的,因为它需要通过筛选识别出那些百万分之一的细胞,就是在其内通过同源重组与由实验者提供的修改过的版本之间交换了一个基因。
1990年代中期,哺乳动物学家在酵母遗传学的观察基础上发现在某个基因位点上引进一个双链断裂可以极大地增强同源重组和由非同源末端接合导致的小缺失的发生频率(综述见Haber,2000和Jasin与Rothstein,2013)。他们意识到,高效基因组编辑的秘诀在于找到一种可以在任何想要的位置制造出双链断裂的可靠手段。最初的普遍策略是使用锌手指核酸酶(ZFNs)——一种由一个锌手指DNA结合域和一个取自限制性内切酶的DNA切割结构域所组成的融合蛋白,它可以结合并切割基因组位点(Bibikova 等,2001)。不久科学家们就在果蝇和小鼠身上证明了ZFNs依靠同源重组对于具体位置上的基因组编辑的用途(Bibikova 等,2003;Porteus和Baltimore,2003)。
到2005年,桑加莫生物科学公司(Sangamo Biosciensces)的研究小组报告了针对在人类细胞系上造成重度联合免疫综合症的基因的突变进行了成功的修正(Urnov 等,2005)。然而,塑造能够可靠辨认具体位置的ZFNs的过程是缓慢而费力的。更好的方法则出现在2009年下半年,两个研究小组描述了一组来自于植物病原体黄单胞菌(Xanthomonas)(Boch 等,2009;Moscou和Bogdanove,2009),叫做TALEs的特殊转录激活蛋白,TALE使用一个特定的模块密码来瞄准具体的DNA序列。但是,这一方法还是需要可观的工作量,因为针对每个目标都要求设计一个新的蛋白。
自从Marraffini和Sontheimer的2008年论文证明了CRISPR是一种可设计的限制性内切酶以来,研究人员已经意识到如果它可以在哺乳动物细胞内发挥作用的话,CRISPR也许就能为具体基因位点的切割和编辑提供强大的工具。但是,这个“如果”是非常关键的。相对于微生物,哺乳动物细胞拥有不同的内部环境,它们的基因要大1000倍,位置在细胞核内,并且嵌在一个精致复杂的染色质结构内。转入诸如自我剪接II类内含子的其它简单微生物系统已经失败了,而使用核酸来瞄准基因位点的尝试也问题重重。CRISPR究竟能否被设计构造成为一种用于编辑人类基因组的强力工具?直到2012年9月,专家们是持怀疑态度的(Barrangou,2012;Carroll, 2012)。
张锋11岁时从中国石家庄搬到爱荷华州的得梅因(Des Moines, Iowa)。他在一次周六兴趣课程中被分子生物学深深吸引,在16岁时就在一家当地的基因疗法实验室每周工作20小时。
在哈佛念本科的时候,他因为室友患了重度抑郁症的事而开始对大脑发生兴趣,后来他到斯坦福跟随神经生物学家兼精神病学家Karl Deisseroth攻读博士学位,期间他们(联同Edward Boyden)发展出了光遗传学(optogentics),这是一项革命性的技术,依靠此技术可以通过光束来触发携带微生物光敏感通道蛋白的神经元。在波士顿身为独立研究员期间(先后在哈佛和麻省理工学院的脑及认知科学系还有博德研究所[Broad Institute]作初级研究员),张致力于进一步扩展研究神经生物学的分子工具。
在开发出一种使用光来激活基因表达(通过把一个DNA结合结构域和一个转录激活结构域同两个在光条件下相互结合的植物蛋白偶联)的方法之后,他开始探索一种用来设计转录因子的普遍方法。在TALEs被解码之后,张与他的合作者Paola Arlotta和George Church(此外还有一个来自桑加莫生物科学公司的研究小组)都成功地将TALEs改造用于哺乳动物,使得精准激活、抑制和编辑基因成为可能(张等,2011;Miller等,2011)。然而,张锋并没有停止寻找更好的方法。
2011年2月张听了哈佛的微生物学家Michael Gilmore关于CRISPR的报告就立刻被它吸引住了。他次日飞赴迈阿密参加一个学术会议,却闷在酒店房间里一头扎进CRISPR的文献资料。回来之后,他就着手创造嗜热链球菌的Cas9版本用于人类细胞(用优化密码子和一个核定位信号)。到2011年4月,他已经发现,通过表达Cas9基因和一个设计过的靶向携带荧光素酶的质粒的CRISPR RNA,他可以在人胚胎肾细胞(HEK)内降低荧光度。但是,效果还是不尽如意。
接下来的一年间张一直在优化这个系统。他探索着可以增加进入细胞核的Cas9比例的方法。当他发现嗜热链球菌Cas9在细胞核内并非平均分布(它在核仁处聚集),他就测试其他的选择,发现化脓性链球菌(S. pyogenes)的Cas9分布地更均匀。他发现哺乳动物细胞虽然缺乏细菌的RNaseIII,但是仍然可以处理crRNA,尽管是用和在细菌里不同的方式。他测试了tracrRNA的各种同工型(isoform)来造鉴定在人类细胞内保持稳定的那一个。
到了2012年的中期,他已经获得了一个由三部分组成的强有效的系统,包含来自化脓性链球菌和嗜热链球菌两者之一的Cas9、tracerRNA和CRISPR阵列。以人和小鼠基因的16个位置为靶向,他证明了,高效而准确地改变基因是可能的——通过非同源重组的末端接合制造缺失和通过与修复模版进行同源重组来实现插入新序列。此外,多重基因可以通过设计CRISPR阵列,用彼此匹配的间隔来同时编辑。当Charpentier和Doudna的论文在那年初夏发表之时,他还用在她们的实验里描述的短sgRNA融合来测试一个两部分组成的系统。
在体内环境下这一融合的效果很差,仅仅低效地切割了位点的一小部分,但是,他发现一个复原了3‘端发夹结构的全长融合可以解决问题(Cong 等,2013;Zhang,2012)。(张之后很快继续证明了CRISPR比原想的更加功能多样:它可以被用于在数周内创造遗传疾病和体细胞癌的复杂小鼠模型,也可以用于实施全基因组筛选来寻找某一生理过程中的必须基因——它的精确度还可以通过降低脱靶切割来提高。
他和曾与van der Oost共事的计算机生物学家Koonin还将发现新的第2类CRISPR系统,包括那个与Cas9切割方式不同而且仅需要crRNA却不需要tracrRNA的核酸酶系统[Zetsche 等,2015]。)张于2012年10月5日提交了报告哺乳动物基因组编辑的论文,发表在2013年1月3日的《科学》上(Cong 等,2013);这篇论文将成为该领域内被引用次数最多的论文,而他的试剂由一家非营利性组织Addgene分发,在之后的3年间接受到25000次申请。

清楚张的研究,同时受到Charpentier和Doudna的启发,Church着手在哺乳动物细胞内测试crRNA-tracer融合。和张一样,他也发现短融合在体内环境是无效的,而全长融合则效果好。他瞄准7个位置,证明了非同源重组末端接合和同源重组的作用。他的论文和张的论文是相继发表的(Mali 等,2013)。(Church和其他研究者都在不久之后使用CRISPR来提升“基因驱动力”(gene drive)——合成基因在自然群体中可以迅速扩散,从而引发将其应用于控制携带疟疾的蚊子的惊喜和对于破坏生态系统的担忧。他还将寻求使用CRISPR在猪的基因组里灭活逆转录病毒来推动猪的器官移植给人。)
梅斯医学注:文章的共同第一作者就是天才少女杨璐菡,目前已在国内创办基因编辑公司,进行器官移植探索。总之,基因编辑的早期英雄,不仅在科研上,也在商业上进行率先探索。
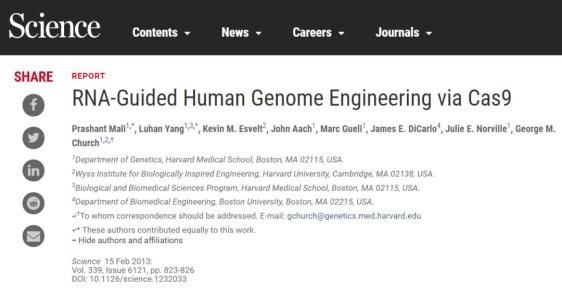
曾从事ZFNs和TALEs基因组编辑的韩国首尔国立大学(Seoul National University)的Jin-Soo Kim 教授报道了在两个位点进行切割(Cho 等,2013)。在以上两个例子中,切割都是低效的,因为sgRNA缺乏tracerRNA的关键性3‘端发夹结构。作为使用ZFNs和TALEs进行基因组编辑的领军人物,哈佛大学教授Keith Joung 则更进了一步。使用由他的合作者Church提供的全长sgRNA结构,Joung通过在斑马鱼的实验证实CRISPR可以在生殖系上有效地制造缺失(Hwang 等,2013)。这些短论文提交于2012年末,在张和Church的论文2013年1月初发表不久之后被接收,于1月末在线发表。
CRISPR迅速蹿红
2013年初,用谷歌搜索“CRISPR”开始井喷,这一趋势至今未减弱。在一年之内,研究人员已经报告了在多种生物内使用基于CRISPR的基因组编辑,包括酵母、线虫、果蝇、斑马鱼、小鼠和猴子。科学和商业上对其在人类医疗和农业生产的潜在应用的兴趣也开始上升,同时这个技术可能被用来培育设计婴儿的前景也引发了社会关注。
CRISPR的早期开拓者并未停止拓展边界,但是他们不再孤单。全世界的科学家都涌入了这一领域,他们是一批新的英雄队伍,他们进一步地阐明CRISPR的生物学,推进和扩展了基因编组辑技术,并且将它运用于广泛范围的生物学问题。在这篇文章的范围内公正描述他们的贡献是不能的;读者可以参考最近的综述(Barrangou和Marraffini,2014;Hsu 等,2014;van der Oost 等,2014; Sander和Joung,2014;Jiang和Marraffini,2015;Sternberg和Doudna,2015;Wright 等,2016)。
发现20年前的一处西班牙盐沼而曾经不为人知的微生物学系统如今却成了科学期刊特刊、《纽约时报》的头条、生物科技创业公司和国际伦理学峰会的焦点。CRISPR的时代已经到来了。
CRISPR的启迪
CRISPR的故事充满了关于产生科学进步的人文环境的启迪,与学术资助机构、一般公众和踌躇满志的研究人员相关。
其中最重要的一点是医学突破往往来源意想不到的地方。CRISPR的早期英雄们都不是专门研究编辑人类基因的,甚至都是不是研究人类疾病的。他们的动机混合了个人兴趣(弄清楚耐盐细菌内奇怪的重复序列)、军事上的特殊需要(防范生物战争)和工业应用(提高酸奶产量)。
这段历史还说明了基于大数据的“无假设驱动”式发现在生物学研究中愈发重要的位置。CRISPR基因位点,它的生物学功能和tracrRNA的发现都不是来自实验台上的实验,而是来自对大范围、常常是公开的基因数据库的生物信息学的开放式探索。“假设驱动”科学当然依旧是基本的研究方法,但是21世纪将会看到更多的是两种科学研究方法的结合。
如此众多的CRISPR英雄们都在科学生涯的开头(包括Mojica,Marraffini,Charpentier, Vogel还有张)于30岁之前就做出了他们各自的重要成果是很有启发意义的。年轻常常意味着愿意在未知的方向和看似无人问津的问题上冒险——和想要获得成功的动力。对于如今这个首次获得NIH资助的年龄已经升至42岁的时代而言,这是个重要的提醒。
还值得注意的是,他们中的许多人都是在可能被一些人看作远离科学研究的常规渠道的地方做出标志性工作的(西班牙的阿利坎特;法国的国防部;丹尼斯科的公司实验室;Vilnius在立陶宛)。此外,他们的重要论文都被一流期刊拒绝了——在很久的延迟之后得以发表在并不令人瞩目的位置。这些遭遇恐怕并非偶然:这些研究地点可能给予了从事不那么热门选题研究的更大自由,但是对于如何克服期刊和评审人的怀疑与不信任态度所给予的支持帮助却是较少的。
文章引起巨大争议
Eric Lander的文章发表后,一些批评者对此提出了激烈的反对意见。问题在于,Broad研究所作为共同专利权人,卷入了美国专利和商标局(USPTO)正在展开调查的一场知识产权战之中。并且Lander的Cell文章没有披露潜在的利益冲突。
加州大学伯克利分校的Jennifer Doudna与德国Helmholtz感染研究中心的Emmanuelle Charpentier一起,当前正与Broad研究所的张锋及同事们陷入一场专利纠纷中。1月17 日,Doudna 在PubMed Commons网站上发表评论称Lander的描述“与事实不符”。Doudna指出,Lander并未核对针对她的实验室以及实验室与其他研究人员交往的描述,且在文章发表前也未征得她的同意。
在写给《科学家》(The Scientist)杂志的一份声明中,Lander说他确实“向Cell杂志透露了现实与可感知的冲突,”包括他的机构拥有的一些CRISPR专利及专利申请。Lander还说,他在12月中旬向Doudna发出了电子邮件,请求她核查将在他的观点文章中发布的材料。
Lander写道:“她确认了有关她的个人背景信息,但表示不想以任何方式评论有关CRISPR技术开发的历史陈述。在撰写这篇文章的过程中,我收到了世界各地十多名科学家提供的关于CRISPR开发的信息。令人遗憾地是,Doudna是其中唯一一个表示拒绝的。尽管如此,我完全尊重她的决定,没有分享她的观点。我也明白将会有不同的观点。”
Doudna告诉The Scientist,Lander确实在12月18日联系了她,但他只共享了文章的一部分。Doudna在一封电 子邮件中写道:“他拒绝与我一同分享许多有关我实验室研究的部分。在发表之前我从未看到过完整的文章,我有电子邮件通信证实这一点。Lander应该写出为他提供信息的其他科学家的名字。”
任职于哈佛大学和Broad研究所,一直与张锋和其他研究人员合作开展CRISPR研究的George Church是其中的一位科学家。此前Church曾对外表达,世人不应CRISPR技术仅归功于张锋等三人,他也是其中的重要贡献者之一。Church在给The Scientist的一封电 子邮件中写道:“Eric Lander在12月14日问了我一些非常专业的问题,我提供了事实核查。他在1月13日发给我一份预印件。我立即寄给他一份有关事实错误的清单,但其中没有一个错误被纠正过来。”
PubPeer网站在这篇论文发布后不久便出现了一些匿名评论。一位用户写道:“这非常的可疑。在这场经常被称作为生物技术史上最大的专利战中,我们是中立的一方。在这场争斗中,Cell居然允许卷入专利战中的一家机构的领导者——Eric Lander决定谁应该突显为这些研究发现的英雄。”另一位用户则写道:“这篇文章在毫不掩饰地为张锋宣称所有权,而Broad研究所冠冕堂皇地从中获益。”
加州大学伯克利分校Michael Eisen在他的网站上谈论CRISPR和相关的专利纠纷时透露了他的潜在冲突,在推特(Twitter)上Eisen提出了针对这篇文章的几点反对意见。他在其中写道:“Lander最具破坏性的错误描述 是,他所有的‘#CRISPR英雄’都是项目负责人(PIs)——那学生和博士后呢?”
在发给The Scientist的一份声明中,Cell出版社发言人Joseph Caputo重申,Lander提供了一份利益冲突(COI)声明,尽管Lander声明了有机构冲突,但没有声明有个人的经济利益冲突。Cell出版社的COI政策不适 用于机构冲突,因此Cell没有发表声明。Caputo在一封电子邮件中写道:“我们当前正在评估我们的COI政 策,以确定我们是否应该进一步扩展它纳入机构利益冲突。我无法对是否会改变Lander的文章做出评论。”
2016-2020年,基因编辑飞速发展
前面的综述发表于2016年,总结了基因编辑技术,尤其是CRISPR技术的历史,事实上,在2016年以后这5年,基因编辑技术又有突飞猛进的发展。这个发展源于经典的基因编辑的脱靶问题。
虽然,基因编辑具有强大的DNA编辑的功能,但是脱靶的现象令人担忧,因此,寻找新型基因编辑系统,变成了关键。在最近5年,基因编辑的技术又出现许多大神级人物,将基因编辑改造得更进一步,越少的脱靶效应,更精准的编辑,甚至进行单碱基编辑,以及推向临床应用。
2020年6月22日,博德研究所David Liu团队在Nature Biotechnology 在线发表题为“Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors”的综述文章,该综述首先描述已表征的Cas9和Cas12核酸酶的天然变异体,并详细介绍具有扩大的靶向范围和特异性的Cas9和Cas12核酸酶变异体的开发。
接下来,该综述讨论碱基编辑器的开发和应用,这些编辑器可精确安装点突变而无需双链DNA断裂(DSB)或供体DNA模板。最后,该综述总结了新兴的CRISPR–Cas基因组编辑工具,包括介导大片段DNA重排的Cas转座子和重组酶,以及主要编辑器,它们以取代原始DNA序列的方式直接将编辑后的序列复制到目标DNA位点。

接下来,该综述讨论碱基编辑器的开发和应用,这些编辑器可精确安装点突变而无需双链DNA断裂(DSB)或供体DNA模板。最后,该综述总结了新兴的CRISPR–Cas基因组编辑工具,包括介导大片段DNA重排的Cas转座子和重组酶,以及主要编辑器,它们以取代原始DNA序列的方式直接将编辑后的序列复制到目标DNA位点。
单碱基编辑器介导的基因组编辑
单碱基编辑器无需诱导DSBs和补充DNA模板的条件下便可实现靶位点的精准点突变,且该过程不依赖于HDR修复途径。目前可用的单碱基编辑器是将活性受损的CRISPR-Cas核酸酶(无诱导DSBs的能力)与单链DNA脱氨基酶融合而得,特定条件下,还需融合可调控DNA修复机制的蛋白。
现有的单碱基编辑器有两大类:一类是融合胞苷脱氨基酶的胞嘧啶碱基编辑器(CBEs),可催化C·G--T·A碱基对的转换;另一类则是融合优化过TadA*脱氧腺苷脱氨基酶的腺嘌呤碱基编辑器(ABEs),可实现A·T--G·C碱基对的转换。CBEs和ABEs可有效介导四种碱基转换突变(C-T,A-G,T-C和G-A),这类突变约占目前人类致病变异的30%。目前单碱基编辑器被广泛用于多种细胞系和模式生物中,并用于多种人类遗传病动物模型的构建和治疗
单碱基编辑器借助于Cas蛋白实现DNA定位,进而实现靶位点处特定活性窗口内目标核苷酸的精准编辑。因此,PAM序列对单碱基编辑器的编辑范围有重要影响。为此早期研究者致力于使用可识别不同PAM的Cas同源蛋白和突变体以拓展单碱基编辑器的编辑范围。
此外,通过将脱氨基酶与不同的Cas同源蛋白融合,研究者可获得小型化的单碱基编辑器以用于AAV病毒的包装和递送。此外,研究者还利用内含肽对单碱基编辑系统加以拆分,从而可通过双AAV系统实现高效的在体单碱基编辑。
考虑到各类基因编辑工具的靶位点编辑效率因位点、细胞类型和细胞状态而异,开发更为多样化的单碱基编辑工具对单碱基编辑的未来应用有重要意义。
ABE单碱基编辑器的新风险——介导胞嘧啶的脱氨基化
2017年10月25日,来自哈佛大学化学与化学生物学系、HHMI以及Broad Institute的David Liu教授实验室在Nature杂志上以长文形式(Article)发表了题为“Programmable base editing of A•T to G•C in genomic DNA without DNA ”的突破性成果,报道了一种新型腺嘌呤碱基编辑器 (Adenine Base Editor,ABE),它可以将A•T碱基对转换成G•C碱基对,加之此前报道的将G•C碱基对转换成T•A 碱基对的成果,该技术首次实现了不依赖于DNA断裂而能够将DNA四种碱基A、T、G、C进行替换的新型基因编辑技术。
这一突破性技术成果在未来将有望更广泛用于纠正某些引起遗传疾病的单碱基突变,摆脱了以往仅限于将G•C碱基对转换成T•A 碱基对的束缚。可实现A·T--G·C碱基对的转换,这对上述的近半数点突变遗传病的治疗有重要意义,因而在基因治疗领域的应用前景相当广阔。

不过,2019年多项研究指出,ABE系统存在大量的RNA脱靶,好在研究者通过点突变的策略成功的降低了ABE系统在RNA水平的脱靶,改善了ABE系统的安全性。
2019年9月23日,来自韩国汉阳大学化学系的Sangsu Bae实验室与首尔国立大学的Jin-Soo Kim实验室合作在Nature Biotechnology杂志上发表了题为Adenine base editors catalyze cytosine conversions in human cells的论文。文章对ABE系统介导胞嘧啶(C)编辑的能力进行了系统分析。
文章证实ABE系统编辑胞嘧啶(C)的能力并非偶然,它能编辑位于sgRNA位点5-7位(PAM为21-23位)的TC*N序列中的胞嘧啶(C),该能力独立于其介导A·T--G·C碱基对转换的能力,且最高效率可达11.2%。
2020年5月11日,华东师范大学李大力课题组在Nature cell Biology发表了题为Increasing the efficiency and targeting range of cytidine base editors through fusion of a single-strand DNA binding protein domain的研究论文,报道了一系列超高活性的胞嘧啶碱基编辑器(hyCBE),证实其提高编辑效率、扩展编辑窗口的同时保持高效精准的工作性能。
不过,基于低活性胞嘧啶脱氨酶所构建的胞嘧啶碱基编辑器,可能无法有效介导在组织器官或者体细胞(如原代血细胞)中的基因组靶向碱基编辑。因此,如何发展新策略构建低脱靶率的高效碱基编辑系统将成为碱基编辑领域未来发展的难点和突破点。
转座酶和重组酶介导的基因编辑
活细胞基因组中的定向整合一直极具挑战性。虽然核酸酶和单切口酶介导的HDR可实现基因组中的定向整合,但仅局限于可分裂的细胞。近年来研究者发现自然界中存在CRISPR转座酶系统可实现细菌基因组中的定向整合;研究者还将Cas蛋白与转座酶融合,同样可实现细菌基因组中的定向整合。此外,研究者还将Cas蛋白与重组酶融合,可在人源细胞中实现目标DNA片段的删除。
CRISPR转座酶系统 对CRISPR系统多样性的探索让研究者发现了可在细菌中适用的CRISPR转座酶系统
dCas-转座酶及重组酶融合系统 将转座酶或重组酶与dCas9融合或有助于定向转座和重组。目前而言,dCas-转座酶融合系统可在大肠杆菌中发挥一定效果;而dCas-重组酶融合系统则能在哺乳动物细胞中实现目标DNA片段的删除,当然,其编辑效率不尽如人意,且适用范围相当有限。
CRISPR转座酶及重组酶系统的未来发展 CRISPR转座酶及重组酶系统依然处于起步阶段,虽然CRISPR转座酶系统在体外和细菌系统中的效果令人欣喜,但其仍难以在哺乳动物细胞中发挥功能。因此,深入探究CRISPR转座酶系统的作用机制并开发真核细胞可用的定向转座系统将是基因编辑领域的重大突破。
引导编辑(Prime editing)
CRISPR-Cas核酸酶可诱导DSBs,在末端连接修复途径下可实现DNA序列的删减和破坏;而在HDR途径下则可实现精准修复,不过常伴有严重的插入缺失脱靶突变。单碱基编辑可在不诱导DSBs的条件下实现精准的单碱基编辑,但只能诱导碱基转换而无法实现碱基颠换。当然,单碱基编辑也无法实现精准的插入缺失编辑。此外,单碱基编辑器常会导致靶位点非目标碱基的脱靶突变,且部分位点由于缺乏合适的PAM序列而难以实现目标碱基的精准编辑。2019年出现的引导编辑有着广谱的编辑能力,理论上可精准诱导所有类型的单碱基突变、小片段插入和缺失突变。
线粒体:基因编辑最后一个角落
《自然》在线发表了关于基因编辑的一篇重要论文。来自华盛顿大学的Joseph Mougous教授与Broad研究所的刘如谦(David Liu)教授展开合作,开发出“第一款实现线粒体DNA精确编辑的分子工具”。
这项研究成果一经发表迅速引起业内的点赞。《自然》《科学》等顶尖学术期刊都特地刊发新闻介绍。研究线粒体疾病的专家评论说,这项研究用“非常巧妙的策略解决了线粒体领域的‘圣杯’问题”。

线粒体内的DNA编码13种蛋白质,虽然不多,但每一种都参与细胞的能量供应。线粒体DNA突变可能导致数十种代谢疾病,包括心肌病、Leber遗传性视神经病变等,而且这些疾病至今没有治愈方法。如果有基因编辑工具可以精确纠正线粒体DNA,无疑将为治疗这类疾病打开大门。
微生物学家Mougous教授与同事们在研究细菌之间如何用毒素相互攻击时,有了一个意外发现。他们找到一种细菌脱氨酶,可以直接修改双链DNA上的碱基,把其中的胞嘧啶(C)转变成尿嘧啶(U),从而杀死其他细菌。过去微生物学家们发现过一些细菌脱氨酶,而它们都是对单链DNA或单链RNA起作用。因此,作用于双链DNA的脱氨酶毒素显得十分异常。
科学家们敏锐地意识到,“这种细菌毒素的特性可以让它与非CRISPR的DNA定位系统配对,”刘如谦教授说道,“使我们最终在生物学的最后一个角落——线粒体DNA中进行精确的基因组编辑。”
正如《自然》评论的,实验性地改变线粒体基因组,可以帮助我们“更好地了解线粒体DNA突变在复杂疾病、癌症和衰老相关的细胞功能障碍中的作用”,同时这一研究“为发展线粒体疾病基因疗法做出了关键进展“,让我们看到了一个能够有效治疗线粒体疾病的未来。我们期待这一天的早日到来。
基因编辑,极富应用前途,它真能改变未来医学。
基因编辑将来在基因治疗方面,应该成为主流技术,其想象空间,是当前任何一项医疗技术难以比美的。
(1)深度学习模型结合基因编辑
在David R. Liu团队综述发表前10天,即2020年6月12日,博德研究所David R. Liu团队在Cell 在线发表题为“Determinants of Base Editing Outcomes from Target Library Analysis and Machine Learning”的研究论文,该研究在哺乳动物细胞中38,538个基因组整合靶标上表征了11个胞嘧啶和腺嘌呤碱基编辑器(CBE和ABE)的序列-活性关系,并使用所得结果训练了BE-Hive,这是一种机器学习模型,可准确预测碱基编辑基因型结果(R ≈0.9)和效率(R≈0.7)。
研究人员以≥90%的准确度纠正了3388个与疾病相关的SNV,其中包括675个等位基因,其“旁观者”核苷酸被BE-Hive正确预测,因此无法编辑。该研究发现了以前无法预测的C-to-G或C-to-A编辑的决定因素,并利用这些发现以≥90%的准确性纠正了174个病原性SNV的编码序列。最后,该研究利用BE-Hive的见识来设计新颖的CBE变体,以调节编辑结果。这些发现启发了碱基编辑,实现了以前难以处理的目标的编辑,并为新的基础编辑器提供了改进的编辑功能。
基因组编辑实验的目的是在细胞基因组的天然背景下,将目标DNA序列转换为新的所需DNA序列。在大多数情况下,期望以高收率获得单个序列产物,但是取决于应用,所编辑序列之间的异质性可能是可接受的。在设计实现预期序列转化的策略时,有必要考虑几个因素。
所需的编辑类型决定了适合的编辑器类别。常见的所需针对性编辑包括以下内容:(i)DNA碱基对的转换(即点突变),(ii)DNA碱基对的缺失,(iii)DNA碱基对的插入或(iv)以上联合的更改(包括DNA碱基对的替换)。不同类别的CRISPR–Cas编辑器介导这些类型的变化。也可能需要其他改变,例如DNA片段的倒置或染色体易位。可以根据序列改变的大小以及所需的效率和产物纯度进一步对所需编辑进行分类。
当前的CRISPR-Cas蛋白库与目标DNA序列结合的能力是第二个主要考虑因素。迄今为止,所有被DNA靶向的CRISPR-Cas系统的靶向都要求在靶DNA位置附近出现一个短序列,称为原间隔子基序或PAM。天然存在的CRISPR系统提供了多种PAM选项,并且开发出了更多经过工程改造的变体,它们具有更宽泛或改变的PAM兼容性。特定工具的最佳选择必须考虑到靶位点上PAM序列的可用性,同时要认识到并非所有基于CRISPR的工具都与所有PAM变异Cas蛋白模块兼容。大多数CRISPR编辑方法还要求PAM和所需编辑位置之间的核苷酸数在一定范围内。
最后,CRISPR–Cas工具的最佳选择和整体编辑策略也将取决于预期的应用。诸如细胞类型(例如细菌,酵母,哺乳动物癌细胞系或哺乳动物有丝分裂后细胞),细胞环境(例如细胞培养物,类器官或体内),试剂形式(例如质粒DNA,核糖核蛋白(RNP)复合物,mRNA或病毒载体等因素)和递送方法(例如,脂质介导,电穿孔,核转染或病毒感染)各自施加了不同的限制,以及对不良基因组修饰事件的不同倾向。
(2)移植领域,3.0版猪问世
在移植领域,在天才少女杨璐菡领衔的国际团队研究下,十几只经过CRISPR基因编辑的“猪3.0”成功问世,这些小猪具备去除猪内源性逆转录病毒及增强异种器官的免疫相容性两个特性,不仅安全且和人有更好的免疫兼容性,有望用于临床的异种器官移植雏形。该研究于2020年9月21日发表在《自然·生物医学工程》杂志上。

(3)单基因疾病最被看好
单基因疾病毫无疑问是基因编辑的首选临床应用对象。几位基因编辑大神都成立相应的公司,主攻方向也是单基因遗传病为主。确实,这方面突破很迅速,只要解决脱靶问题,就能很快应用于临床。β-地中海贫血和镰刀状贫血更是关注的重点。
2019年3月25日,一项最新的研究表明,通过CRISPR/Cas9基因编辑技术编辑β-地中海贫血和镰刀状贫血患者造血干细胞中的BCL11A增强子位点,并进行自体造血干细胞移植,可以使得体内分化产生具有高比例HbF及正常功能血红蛋白的红细胞,表明CRISPR/Cas9介导的基因编辑技术有望彻底根治由β-globin珠蛋白突变引发的系列遗传疾病。
这项研究以 Highly efficient therapeutic gene editing of human hematopoietic stem cells 为题在线发表于最新的国际顶级医学期刊Nature Medicine 杂志上。该论文的通讯作者为:Daniel E. Bauer(波士顿儿童医院);第一作者为:吴宇轩(华东师范大学生命科学学院)、曾静(波士顿儿童医院)。

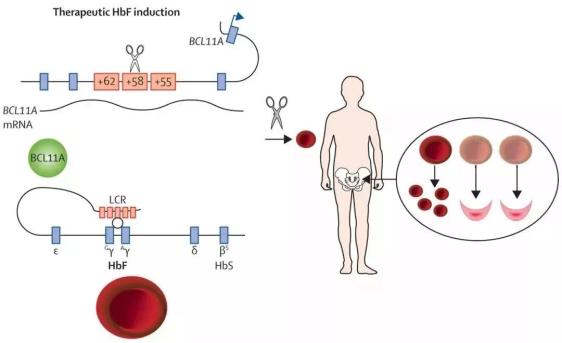
基于上述机理,最新研究首先通过比较不同的sgRNA优化编辑系统,实现了CRISPR/Cas9核糖核蛋白(RNP)在人CD34+造血干/祖细胞中的高效基因编辑。接着,将编辑过后的CD34+细胞体外红系分化后,发现BCL11A的表达水平显著降低,同时HbF的含量得到极大提升。
先天性黑蒙症是导致儿童失明的最常见原因之一,据美国国立卫生研究院的数据,每10万名新生儿中约有2至3名患有此病。该疾病是发生最早和最严重的遗传性视网膜病变,患儿通常在出生时或出生后一年内双眼视锥细胞功能完全丧失,导致先天性失明。
新的疗法将使用CRISPR来纠正基因突变。CRISPR是一种基因编辑工具,可以让研究人员精确地编辑特定位置的DNA。根据Editas Medicine公司的一份声明,研究人员将使用一种注射剂,直接在感光细胞中进行这种疗法。Editas Medicine公司正在与艾尔建(Allergan)公司一起开展这项研究,该试验将纳入18名患者,包括儿童(3岁及以上)和成人,在这项新的研究中,儿童和成人的DNA编辑不能遗传给他们的后代。
(4)纳米递送载体,脱离病毒载体,让基因编辑离临床更近
2020年4月份Nature Nanotechnology上,Qiang Cheng等人提出了一种称为选择性器官靶向(SORT)的方法,通过生物工程将含有核酸疗法的LNPs诱导肝脏、脾脏和肺特异性基因调控。LNP系统通常由磷脂、胆固醇、聚乙二醇(PEG)脂质和可电离的阳离子脂质组成。每个LNP成分及其摩尔比都已被优化,以确保高效的核酸递送到肝细胞。特别是,合理设计和筛选可电离的阳离子脂质(或类脂质)文库对于实现临床应用相关剂量的肝基因沉默至关重要。
最有效的电离阳离子脂质,其特征是明显的pKa值在6.2和6.5之间,具有三种功能:(1)在LNP生产期间,酸性pH值下质子化的叔胺脂质头部促进其与带负电荷的核酸结合,(2)在生理的pH值,附近无电荷的脂质确保净中性表面电荷,以减少免疫反应和循环时间延长和(3)细胞摄取后进入酸化的内体中,正电荷脂质促进膜融合和有效的胞质递送。
在这种SORT方法中,作者添加五分之一脂质成分来通过静脉注射递送功能信使核糖核酸或基因编辑复合物道特定组织。使用一个快速的混合过程,这种SORT脂质 (永久的阳离子和阴离子或可质子化的阳离子)和不同摩尔比的其他材料生成一个脂质体文库,包载mRNA编码荧光素酶进行体内筛选。通过增加永久性阳离子脂质1,2-二烯烃-3-三甲氨基丙烷(DOTAP)的摩尔百分比(0-100%),可使荧光素酶的表达在静脉注射后从肝脏转移到脾脏和肺。LNPs中加入10-40%的永久性阴离子脂质1,2-二烯醇化酶-sn-甘油-3-磷酸(18PA)可在脾脏中特异性表达荧光素酶。加入20%的其他可电离的阳离子脂类,如1,2-二烯酰-3-二甲基氨基甲烷-丙烷(DODAP),不会改变生物分布,但增加了mRNA对肝脏的递送。
该方法的普遍性通过将LNP与其他永久性带电或可电离的阳离子脂类功能化而得到证实,这导致了器官表达的类似变化,且表现出与脂类相关的方式。值得注意的是,SORT-LNP的疗效离不开可电离的阳离子脂质包合。给予肺、脾脏和肝脏特异性mRNA SORT-LNP的筛选结果表明,基于荧光素酶的筛选可产生持续的治疗性蛋白,且无明显毒性。重要的是,SORT方法还允许调节LNPs组织特异性荧光素酶的表达,使其与临床批准的Onpattro配方相同。
Cheng等人也报道了使用SORT-LNPs进行组织特异性CRISPR/Cas基因编辑。相对于治疗性基因沉默或表达,基于CRISPR/Cas的基因编辑需要(至少)两个组成部分:引导RNA (guide RNA, gRNA)识别目标DNA和Cas核酸酶进行双链断裂。作者生成了包含gRNA和Cas9 mRNA或gRNA/Cas9核糖核酸蛋白复合物的SORT-LNPs,并在肝外组织中进行了基因编辑。在红色荧光蛋白tdTomato报告小鼠中,不同种类的LNP剂型可选择性诱导肝、肺、脾特异性基因编辑。此外,作者还展示了内源性靶基因(PTEN)和治疗性靶基因(PCSK9)的组织特异性编辑。
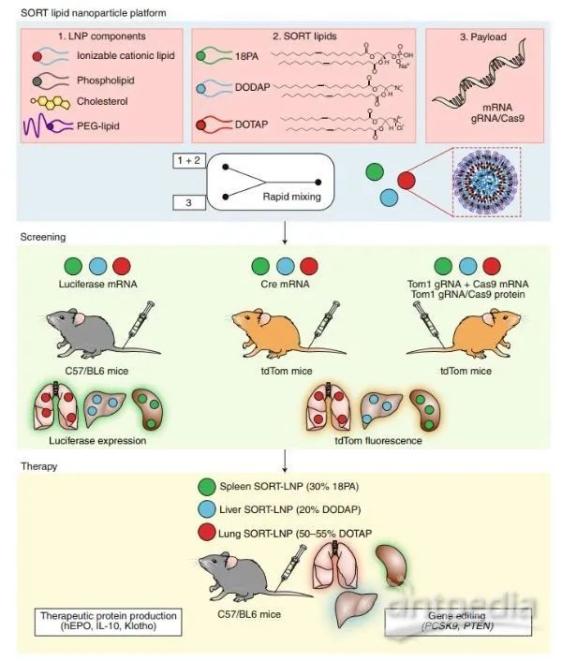
Cheng等人的研究的主要创新点是在复杂的报告模型中采用了系统的LNP筛选方法。Sago等人报道了一种类似的方法,称为纳米颗粒传递的快速识别(FIND)。他们的策略包括筛选数百个LNPs,以确定使用合理设计的DNA条形码将功能性mRNA(和其他RNA疗法)递送到各种组织的有效配方。
南京大学现代工程与应用科学学院教授宋玉君介绍,1.0版的基因剪刀,在“剪辑”时通常是“由外向内”,2.0版的“基因剪刀”则是“内外呼应”。
针对基因剪刀的脱靶效应问题,团队进行了长达一年半的实验。根据实验,研究人员新开发的方法采用了一种名叫“上转换纳米粒子”的非病毒载体,这种纳米粒子具有光转化效果,当它们携带“基因剪刀” CRISPR-Cas9进入细胞内部后,在无创的近红外光的照射下,可以发射出紫外光,这些光线就像交通指示灯一样,为“基因剪刀”提供指令,使得Cas9蛋白不再“路痴”,可以准确进入病变组织的细胞核,从而实现精准的基因剪切。宋玉君说,红外光具有强大的组织穿透性,这为在人体深层组织中安全、精准地应用基因编辑技术提供了可能。
(5)基因编辑与细胞治疗,完美搭配
利用CRISPR/Cas9技术改进CAR-T主要集中在以下几个几点:包括通用型CAR-T(UCAR-T)的开发;通过基因编辑技术能够赋予CAR-T技术全新的高效性、稳定性等优点,相信CAR-T与基因编辑合作碰撞出的火花,将会在新一代细胞免疫治疗的领域展现出更惊艳的成绩,以提高CAR-T细胞的增殖能力和存活时间
(6)基因编辑用于肿瘤,永远的热点
在今年初,顶刊《科学》上刊登了癌症免疫疗法的临床试验最新进展,3名难治性癌症患者接受了一种结合基因编辑(CRISPR)技术的新型T细胞疗法的治疗。这是美国首次在癌症患者身上测试基于CRISPR基因编辑手段改造的细胞疗法。编辑后的 T细胞体内存活时间长达9个月,未出现任何严重不良事件,CRISPR用于癌症治疗安全性过关。

2020年4月27日,华西医院肿瘤中心主任卢铀教授主持的全球首个基因编辑技术改造T细胞治疗晚期难治性非小细胞肺癌的临床试验结果,正式发表在了《自然.医学》杂志上。该研究的试验对象是晚期非小细胞肺癌患者,通过使用“魔剪”技术编辑非小细胞肺癌患者T细胞的PD-1基因后再回输到患者体内展开治疗,这是一项临床1期研究成果。
这项临床试验是2016年10月启动的,卢铀教授利用基因编辑技术(CRISPR-Cas9)改造T细胞进而用来抗癌的临床试验,当年的10月28日,第一位肺癌志愿者接受了基因编辑改造后的T细胞回输治疗。

(7)基因编辑的脱靶的评估,是决定基因编辑能否进入临床另一道门槛
2017年11月16日,美国FDA宣布全球著名的斯隆•凯特琳癌症研究中心(Memorial Sloan Kettering Cancer Center,简称MSK)基于二代测序技术的癌症基因检测分析平台MSK-IMPACT™获得批准。这被视为2017年又一重大科学成果,影响深远,将开辟癌症基因检测的学术研究和商业化先河。
AMP最突出的优势是能发现和检测新的融合基因。麻省总院研究人员曾应用 AMP 技术,相继发现ROS1、RET、NTRK1、NTRK2、NTRK 3 等新的基因融合,ROS1 还被证实对克唑替尼敏感。这帮助医生准确诊断ROS1病例(其他方法常常检测为阴性),效果很显著,该成果也发表于2014 年 11 月的《新英格兰医学杂志》上。实际上,除基因融合外,AMP也同时对其他变异类型进行检测,包括点突变、插入和缺失、拷贝数、RNA表达等,几乎覆盖所有突变类型,可以使用 DNA 或 RNA 为模板,能对数个至数百个基因同时进行平行检测。
CRISPR可以对基因组内的任何基因进行修改,能高效地对任何生命体进行基因组“手术”。然而CRISPR基因编辑技术虽功能强大,但隐藏着巨大的安全隐患,因为切割不准确会造成脱靶。在应用上,我们在乎被编辑过的每一个细胞,因为哪怕仅有一个细胞因脱靶导致癌基因被激活或者抑癌基因被敲除,都会对患者的安全埋下隐患。基于AMP技术平台的全基因组无偏寻找CRISPR脱靶技术(GUIDE-Seq)能全面、灵敏地找出CRISPR编辑脱靶位点。AMP技术平台是经过了临床考验的一个平台,并且获得了全美审查最严格的纽约州的认证。因此,基于AMP技术平台而开发的CRISPR脱靶找寻方法可以为各种以CRISPR为基础的细胞治疗的安全把关,包括CAR-T或TCR-T等疗法,将威胁因素找出来,推动CRISPR技术真正走向临床。
总结
自2018年的“基因编辑婴儿”事件以来,围绕着基因编辑的讨论不绝于耳。这项技术的突破,同时也让大家充满了期待与担忧。有人觉得这对于经受巨大精神创伤及抑郁症的人来说,是福音。也有一部分人觉得这就是“潘多拉的魔盒”,一旦打开就无法轻易关上。
基因编辑是划时代技术,是魔剪,但这个是魔是恶魔,还是祛除恶魔,就看人类往哪里走了,无论怎么说,基因编辑都是面向未来的技术,也许有一天,我们会在路上发现很多具有超能力的基因编辑人,也许并不奇怪。
原始出处:
Lander ES. The Heroes of CRISPR.Cell. 2016 Jan 14;164(1-2):18-28. doi: 10.1016/j.cell.2015.12.041.
Anzalone AV, Koblan LW, Liu DR. Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors.Nat Biotechnol. 2020 Jul;38(7):824-844
Mok BY, de Moraes MH, Zeng J, Bosch DE, Kotrys AV, Raguram A, Hsu F, Radey MC, Peterson SB, Mootha VK, Mougous JD, Liu DR. A bacterial cytidine deaminase toxin enables CRISPR-free mitochondrial base editing.Nature. 2020 Jul;583(7817):631-637. DOI: https://doi.org/10.1038/s41586-020-2477-4
文章主要内容来自于BioArt, 知乎,梅斯医学等翻译相关资料。
原标题:《诺奖的背后:基因编辑发展史上的幕后英雄和未来发展,是魔剪还是魔鬼?!》
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2024 上海东方报业有限公司

