- +1
前沿综述:相变怎样塑造生命系统
原创 Vikas Trivedi等 集智俱乐部

导语
相变是各类复杂系统中普遍存在的现象,基于物理系统的相变理论与技术,正在被广泛应用于生命生态和社会经济等各类复杂系统的研究中。生物组织由大量细胞构成,不同物理尺度、时间尺度的生物系统,常常表现为不同的宏观特性,并且会发生类似固体/液体/气体等状态的转变。2022年发表于Nature Communications的综述文章,系统梳理了当前关于生物系统各种(类)相变现象,以及对其发生机制、检测手段、特殊变化的学界最新认识,讨论从微观尺度的大量相互作用到宏观尺度的系统特性的涌现过程。理解生物系统中的相变过程,对于进一步打通物理系统、生命系统边界,提炼共性的复杂系统理论方法,具有启发意义,同时有助于加深对于生物发育、疾病发生、生物-环境关系的认识。本文是对该综述的全文翻译。
研究领域:相变,复杂系统,涌现
Pierre-François Lenne & Vikas Trivedi | 作者
赵雨亭 | 译者
刘培源 | 审校
邓一雪 | 编辑
论文题目:
Sculpting tissues by phase transitions
论文地址:
https://www.nature.com/articles/s41467-022-28151-9
目录
摘要
一、组织形态发生的相变
二、相变的起源:分子、细胞和多细胞
三、原位检测相变的实验方法和挑战
四、超越相变:组织刚度的缓慢变化
五、生物与非生物系统类比的局限及未来方向
摘要
生物系统表现出丰富的状态现象学,它类似于物质的物理状态——固体、液体和气体。不同状态阶段是由更小的组成成分——细胞——之间的相互作用产生,这些相互作用表现出宏观特性,如流动性、刚性和对形状及体积变化的抵抗力。以此角度看,从刚性/固态到流动/流体,或从流动到刚性的转变,可以定义许多生物过程中发生的大部分情况,特别是在早期发育与癌症等疾病的进行期间。此外,集体移动的细胞还可以导致生物系统中的运动相变,类似于多粒子系统,其中粒子可以相互作用并显示以特定速度为特征的亚群。在本文中,我们从理论角度并借助生物系统中的实验证据讨论了生物和惰性物理系统之间类比的相似性和局限性。在理解这种转变时,重要的是要承认生物材料的宏观性质及其修饰是由细胞微观性质之间复杂的相互作用产生的,包括生长或死亡、相邻相互作用和基质的分泌,以及生物系统特有的现象。在体内检测相变在技术上是困难的。我们提出了应对这一挑战的新兴方法,并可能指导我们对生物组织的组织与宏观行为的理解。
一、组织形态发生的相变
生物组织(biological tissues)需要对其组成成分(细胞)进行适当的组织(organization),以维持合理的结构和功能。与任何多组分系统类似,组成组分之间的相互作用在很大程度上决定了整体行为或宏观状态。例如,物质的三种状态——固体、液体和气体,微观层面上其组成粒子之间的不同相互作用,在宏观层面表现为流动性、刚性、对形状和体积变化的抵抗力等特性。
生物系统也显示出这种物质状态的丰富现象学(phenomenology),这些物质状态是由细胞与其邻居和细胞外介质相互作用产生的,细胞外介质通常也由细胞本身产生(图 1)。例如,骨头、软骨或树皮是生物学中常见的固体状材料。类似流体的行为通常在生物组织中更常见,尤其是在动物界。特别是在胚胎形态发生过程中,胚胎的形状变化是由于单个细胞形状变化和细胞拓扑重排之间的相互作用造成的——例如,在原肠胚形成过程中,胚胎组织表现得像不能强烈抵抗形状变化的液体。一些上皮组织表现出流体液相特征,在长程上高度有序,如液晶[1]。在某些情况下,将组织状态视为气体状态是合理的,其中单个细胞之间的相互作用最小,并且它们的运动类似于气体分子的运动(图 1)。另一方面,植物细胞由于来自基于纤维素的细胞壁内的液泡的饱和膨胀压力,在集体(例如植物根部、植物脉管系统)中表现出更“永久”的类固体行为。然而,当细胞随着时间的推移完全被其自身的细胞外基质 (ECM) 或矿物质分泌物(例如甲壳类动物或昆虫壳、骨头或树皮、或软骨)(图 1)。
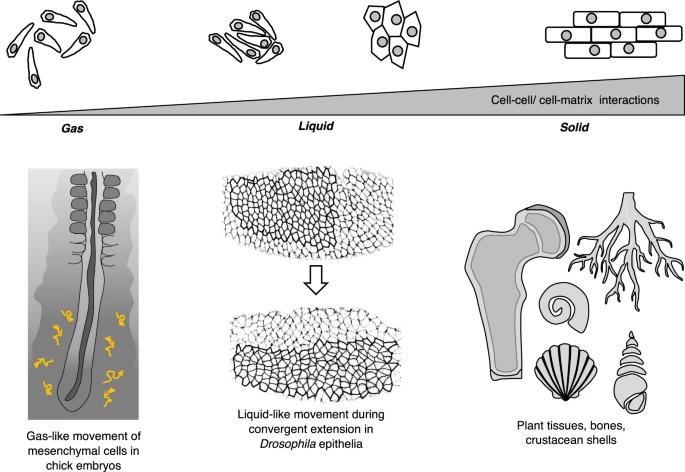
图 1:生物材料显示出物质状态丰富的现象学。与物理物质类似,生物材料可以在群体水平上表现出类似流体和类似固体的特性,这是由细胞与其邻居和细胞外介质的相互作用产生的。组织状态可以是气体状(例如,鸡胚中的间充质细胞[21])、液体状(例如,果蝇原肠胚形成期间的上皮组织[112])或固体状(例如骨头、软骨、树皮)。
特定的分子组成和相邻排列,导致上皮和间充质(mesenchymal)类型的细胞群中出现不同的细胞外特性,从而可以表现得类似固体或液体。通常,在上皮状态下实现细胞的强粘附性和紧密堆积,并且大规模细胞重排表征松散堆积的间充质细胞。然而,尽管在个体水平上有很大的运动,但间充质细胞的集合仍然可以限制在一个小区域内,因此导致该区域的整体结构稳定。最近有研究在斑马鱼中胚层祖区组织逐渐“凝固”的背景下阐明了间充质中这种流体到固体转变的证据,因为细胞进入前体中胚层,在那里它们更加堆积[2]。在这种情况下,细胞密度会影响局部组织,从而控制凝固过程。在细胞密度恒定的汇合组织中,凝固是由细胞间粘附和皮质张力[3]或细胞间和细胞-基质粘附[4]的变化引起的。同样,紧密堆积的上皮细胞可以是“流体状”,其细胞重新排列和/或集体移动[5]。不同的相可以在一个组织中共存;例如,细胞可以形成类似流体的簇,这些簇正在与气体相交换细胞。这种细胞相的共存在体外观察到,对于单个(类气体)细胞从液体或甚至类固体肿瘤发散的肿瘤(类似“蒸发”过程)。表现得像液滴的细胞簇能够从原发性肿瘤迁移并传播,同时保持其上皮特征[7]。
与大多数惰性物理系统不同,生物系统的特点是其组成细胞在不同时间尺度(timescale)上的生长和材料特性的变化。例如,细胞群可以通过改变其微观(细胞内)特性来改变其宏观(细胞上)行为,例如特定的细胞表面或膜蛋白、细胞内细胞骨架的重排、内部细胞器(如植物细胞液泡)的数量、大小和分布的变化。结果是,生物组织的状态通常是短暂的,并且组织可以根据所涉及的分子过程,以快速或缓慢的方式改变其状态。动物细胞中的上皮细胞到间充质细胞的转变就是一个这样的例子,其时间范围可以从几分钟(斑马鱼原肠胚形成)到几个小时(小鼠原肠胚形成)到几天(癌症)。斑马鱼胚层已被证明可以通过调节其细胞间接触的时间尺度,在小时尺度上将其状态从类固体状态变为类流体状态。
似乎这种从刚性到流动状态的转变,可以被定义为生物组织的类固体相(solid-like phase)和类液体相(liquid-like phase),反之亦然,定义了许多生物过程中发生的大部分事情,尤其是在肿瘤发生期间和形态发育期间。值得注意的是,虽然生物组织可以通过分子水平的变化来改变其上述转变状态,但混合的固体和类流体行为也是大多数细胞系统具有粘弹性这一事实的必然结果(框 1)。与惰性粘弹性材料(兼具粘性与弹性)类似,生物组织在较短的时间尺度上也表现出类似固体的行为,在较长的时间尺度上表现出类似粘性流体的行为[9,10]。根据组织的分子组成,这种粘弹性的时间尺度,本身可能会随着时间发生显着变化,并且这种变化可能相当于相关时间尺度上的相变。然而,正如我们后续将描述的,在给定的粘弹性时间尺度上,固体行为到流体行为之间的变化并不是真正的相变(框 1)。因此,在分析生物系统从特定状态(刚性或流体状态)到另一种状态的变化时,考虑该过程的时间尺度至关重要。
相变的标志是系统秩序的变化(框 1)。这种性质变化可以是突然的(即不连续的一级相变)或渐进的(即连续的二级相变)。例如,水从液体到固体的转变会导致水分子作为周期性晶格的组织发生突然变化,而铁磁材料则会显示出内部顺序的逐渐变化。这种突然的或逐渐的变化,通常是由外部条件(控制参数)如温度、压力或密度的渐变引起的。然而,在生物系统中,条件的变化通常是内部的,例如生长速率、细胞分裂、迁移、粘附、排列等。与真正的流体到固体的转变不同,在这种转变中会发生长程结晶顺序的自发涌现,生物系统中的刚性相变的特点是物质在固体和液体中的无序状态持续存在。这种被称为堵塞(jamming,见框 2)的相变已被广泛讨论用于惰性材料,例如泡沫、乳液、颗粒材料和玻璃。受物理学和工程学研究的推动,堵塞已被证明是一种有效的范例,用于定义 2D 和 3D 环境中生物组织中刚性的出现[2,4,11,12.13,14,15,16,17,18,19],其中拥挤、张力驱动刚性和波动减少——所有这三种机制单独或同时——可以阻止细胞运动,从而导致“堵塞的组织”[20]。类似于多粒子系统,其中粒子可以相互作用并表现为具有不同特定速度的粒子群,生物系统也可以在集体移动的细胞中表现出动力学相变(框 1)和堵塞。
在本文中,我们将从理论角度以及生物系统中的实验证据讨论上述相变。特别是,我们将讨论有序系统和无序系统的概念(框 1)在生物学背景下的意义,生物组织中流体到固体转变的主要物理决定因素是什么(类似于压力、密度,尤其是温度),以及哪些标准确定组织中的这种相变。然后,我们将提出在原位检测这种相变的方法。最后,我们还将推测流体/固体转变的分子实现,并批判性地讨论生命系统中的集体效应与常规相变之间类比的机会和局限性。
框1(补充概念)
粘弹性(Viscoelasticity)。粘弹性是由粘性和弹性两个词组成的,表示材料的行为,如聚合物和生物材料,既不作为纯流体也不作为纯固体,但具有粘性和弹性特性。这种材料的机械响应取决于加载或卸载的速度。大多数生物材料在短时间尺度(大约几十秒到几分钟)主要是有弹性的,在长时间尺度(大约几十分钟到几小时)主要是粘性的。
(热力学)相变(Thermodynamic phase transition)。相变描述了物质不同状态之间的转变,如固态、液态或气态。相的特征是物理性质(密度、顺序) ,它在热力学系统中是均匀的。相变通常是由于控制参数(如压力或温度)的变化而引起的。一个典型的例子是,在一个大气压下当温度低于0 °C 时,液态水凝固成结晶冰。在凝固过程中,平移对称性被打破,水分子组成周期性的晶格。在多细胞系统中,除了在特定的器官如昆虫的复眼之外,很少发展出长程的晶体结构,而且与液体/晶体转变的类比似乎不足以描述生命系统。一个更相关的转变类型是堵塞相变(框2),已观察到的大类材料包括颗粒介质、胶体悬浮液、浆体、泡沫和玻璃形成液体。
动力学相变(Kinetic phase transition)。一个部件自组装的系统,可以根据噪声(速度)和密度,经历从部件的随机(无序)运动到对齐(大规模有序)运动的变化。与真正的热力学相变类似,当控制参数通过临界点时,系统动力学行为的这种变化被称为“动力学相变”(也称为聚集相变flocking phase transition)。例如,当细胞密度超过临界值时,以低细胞密度作为个体迁移的细胞(如角质形成细胞),形成连贯的运动细胞群;它们表现出从无序状态到有序状态的转变[33]。在鸟群和鱼群中也观察到类似的转变[104]。
有序系统(Ordered system)。物质的状态表现出不同的结构有序性:晶体固体表现出完美的有序性,它们的组成分子排列成周期性的晶格,而液体则缺乏空间有序性。相变通常以序列变化为特征,例如,液体向固体的转变伴随着结构秩序的变化。然而玻璃是无序的材料,缺乏晶体的周期性,但在机械上表现得像固体。从液体中形成玻璃杯的常见方法是快速冷却,以防止结晶,因此序列重排是不完整的。通过研究密度和方向的空间相关性,可以对有序/无序进行量化。玻璃像液体一样无序,却有像固体一样机械特征(刚性)。
二、相变的起源:分子、细胞和多细胞
惰性物理系统中的相变由有限数量的参数控制,例如温度、体积和应力(框 1,图 2)。鉴于生物系统的复杂性,可以预期参数数量将变得更多,从而将简单的相图转换为复杂的多维相图。潜在的控制参数可以是生化的(pH、O2、信号分子等)或物理的(压力、密度等),从分子到多细胞尺度上起作用。尽管存在这种复杂性,但通过将通常的控制参数替换为潜在的细胞对应物来使标准液固相图适应多细胞系统是很诱人的。例如,在经历类似于堵塞相变的转变的细胞单层中,有人提出用细胞运动来代替温度,用密度代替体积分数,用细胞间粘附的倒数代替应力(图 2)[18]。这些参数具有分子起源,可导致细胞和多细胞水平的变化,从而导致相变。在本节中,我们将讨论相变的实例和来源,并探讨发育和疾病中这种固-液转变的生物学起源。
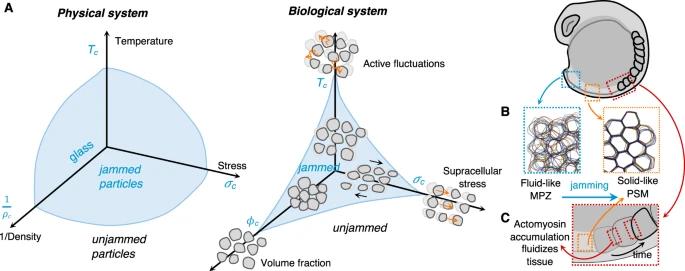
图 2:生物材料中的堵塞相变。A 受温度、压力或密度等外部条件影响的物理系统可以经历从流体到被称为堵塞的刚性相变,而不会自发涌现出长程有序性。生物系统中的类似相图具有三个控制参数:主动波动、细胞外应力和体积分数[2]。B 在斑马鱼的轴向伸长过程中,离开中胚层祖区(MPZ,蓝色)成熟为前胚体中胚层(PSM,橙色)的细胞经历了一个堵塞相变。在 MPZ 内,高细胞间接触长度波动(高有效温度)和更多细胞外空间使组织液状,而前胚体中胚层的细胞重排和细胞混合因细胞外空间较小和细胞间接触波动低而停止(低有效温度)。组织的这种堵塞充当刚性支撑,使组织膨胀(伸长)朝后方向偏置[2,41]。C 年轻体节后边界的F-肌动蛋白和肌球蛋白 II 逐渐积累导致局部张力波动的增加,从而使组织瞬时流体化以进行重塑。一旦由于这种边界张力增加和流化,体细胞结构被物理地夹断,组织就会恢复到其刚性状态,保持体细胞的形状[40]。
细胞运动与重排引起的变化
与惰性系统不同,生命系统中的相变可以由组成成分的动力学和相互作用的变化所触发。细胞运动和细胞重排是这些变化的核心。
细胞运动引起的变化
早期胚胎形态最显著的变化之一是身体形状沿前后轴的延伸 (A-P) 。在脊椎动物中,身体延伸涉及基本的细胞行为包括:细胞迁移、细胞增殖和细胞重排。在雏鸡胚胎中,后伸长主要是由前胚体中胚层 (PSM) 中细胞的增殖和分级随机运动引起的 [21]。从前到后的分级随机运动为胚胎后部的伸长提供了方向偏差。前胚体中胚层的细胞运动分析表明,间充质细胞相对于潜在的细胞外基质表现出布朗运动,表明其具有类似气体的行为(图 1A)。
比较细胞与流体分子,很容易在组织内单细胞扩散和布朗系统中热扩散之间建立类比,从而为细胞运动构建出一个取决于扩散常数的“有效温度”。这个例子表明,在发育过程中,组织可以通过改变其组成细胞的运动特性来改变其流动性。然而,在这个方向上的进一步研究可能有助于我们理解如何将这种生物过程定义为相变现象。
由于主动驱动的细胞重排而发生的变化
流体材料的特点是,颗粒在受到剪切时会相互不可逆地重新排列。在组织中,细胞重排可以由诸如肌动球蛋白网络的收缩等活动过程触发,并且已被证明会导致上皮组织的流化。有助于胚胎成形的细胞重排的一个经典例子是果蝇胚胎的胚带伸长[22, 24]。细胞重排也称为细胞嵌入,涉及由收缩性肌动球蛋白网络驱动的细胞连接的各向异性重塑。它们参与了生物组织的流动性,否则组织会发生弹性变形(图 3B)。
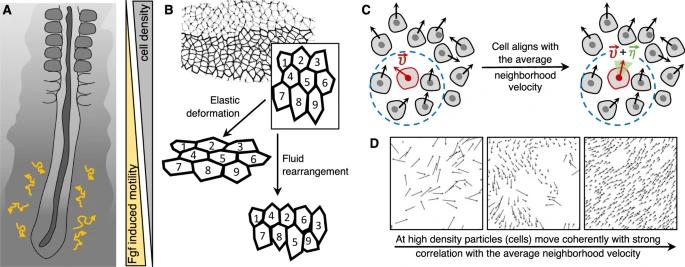
图 3:由细胞运动变化引起的相变。A 在雏鸡前胚体中胚层 (PSM) 中,间充质细胞相对于下面的细胞外基质表现出布朗运动,因此整个组织具有类似液体的行为。当它们在 Fgf 的影响下从后部移动到前部时,它们会经历逐渐的“固化”[21]。B 活跃的细胞重排,由于肌球蛋白依赖的连接重塑,在果蝇胚胎的种带伸长期间,由于邻居交换而呈现组织液态[112]。相反,弹性变形在邻居之间保持相同的配置。C、D 运动细胞的动力学相变标志着自推进粒子系统中自发有序运动的出现。在运动细胞的集合中,每个细胞都可以改变其运动方向,带有一些扰动(由绿色阴影表示)取决于其附近的平均运动方向(由蓝色圆圈表示)。这导致速度的相关性随着粒子移动的密度和噪声的变化而逐渐增加,从而通过旋转对称的自发对称破缺[31,33]。
在雏鸡胚胎中,流动性来自细胞运动的变化,但也是细胞分裂的结果。细胞分裂主动促进了细胞重排:子细胞在分裂后相互分离并重塑它们与附近细胞的连接[25]。这些局部重排是原肠胚成型所必需的,如果细胞分裂被抑制,上皮组织就会变得稳定。在分子水平上,低肌动球蛋白收缩性促进原肠胚形成期间的连接重塑,而在早期阶段,高水平肌动蛋白似乎阻止细胞分裂介导的重排。这些观察结果与预测弹性组织中发生的细胞分裂(和细胞凋亡)引入动态重组的理论模型一致,这种动态重组倾向于具有明确的剪切和体积粘度的类流体行为[26]。
主动驱动的细胞重排导致生物组织尺度的形状变化和流动。越来越多的证据表明,给定细胞重排模式的边界条件会影响宏观变化。在边界为圆形的果蝇生殖器中,上皮细胞的各向异性重排导致单向旋转[27]。这个例子强调了局部力学和形状之间的耦合。
细胞密度引起的变化
细胞密度或等效的体积分数是与相变相关的物理参数,因为它在很大程度上决定了粒子或细胞之间相互作用的频率。在斑马鱼胚胎尾巴的延伸过程中,细胞外空间的体积向后减小并达到一个值,低于该值,细胞材料显示出显著的屈服应力,阻止细胞重排。当自由体积分数低于 0.3628 时,这种流固转变类似于在水性泡沫中发生的情况。Monera等人。表明屈服应力和细胞外液体积的梯度是由细胞粘附蛋白 N-钙粘蛋白控制(尽管蛋白质的浓度本身并不分级)。
密度是与生物组织相关的控制参数吗?上皮组织由密集的细胞组成,这种情况对应的体积分数接近于1。由于在如此拥挤的系统中观察到堵塞和非堵塞状态,因此质疑将密度用作控制参数 是合理的[3,4,16]。早期的上皮机制模型 [29] 确定了机械上不同网络的存在,这些网络让人联想到堵塞和未堵塞的状态:固体状六边形网络和液体状软网络。六边形网络同时具有体积模量和剪切模量。相比之下,软网络的剪切模量则消失了,并且表现得更像一种液体,细胞可以在其中轻松移动和相互穿过。在此类模型中,网络中单元的机械能是三个项的总和:与细胞面积可压缩性相关的项、与归因于细胞刚度的细胞周边弹性相关的项,和与归因于粘附和皮质张力的接触线能量有关的项。细胞形状的变化受到细胞想要获取的优选选周长 P0 和优选面积 A0 参数的限制。在这种转变中,如果细胞的优选周长 P0 相对于优选面积 A0 减小,则组织可以凝固。堵塞/非堵塞相变的控制参数是观察周长与观察面积平方根之间的比率(图 4A)。它被称为形状指数。对于实体组织,形状指数正好是 3.81(这是正五边形的周长与表面积之比)。如果该指数超过 3.81,则组织更像液体。这一预测是在人类患者肺部的上皮单层培养物中实现的,这些培养物表现出堵塞相变。与来自非哮喘患者供体的细胞相比,在来自哮喘供体的细胞培养物中,堵塞相变被延迟。这表明维持上皮细胞的流动性可能是上皮细胞发病的原因之一,而堵塞可能是上皮细胞发挥屏障作用的一种功能状态。最近研究在果蝇生殖带上皮细胞中的工作评估了细胞堆积对形状指数的影响,并表明它实际上可以在 3.72 和至少 3.9 之间变化,具体取决于紧密堆积的上皮细胞中存在多少五边形和多少多重顶点[30]。
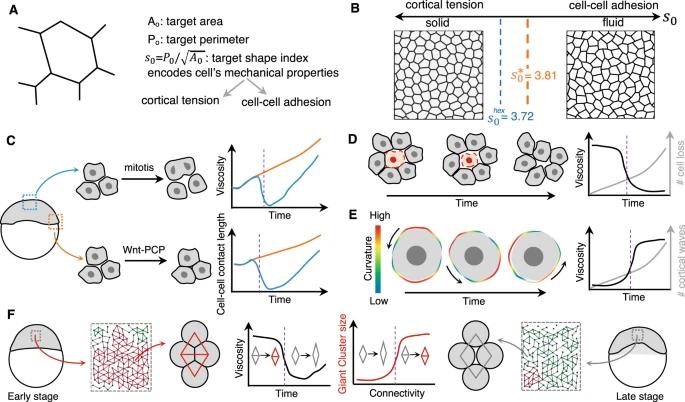
图 4:细胞接触动态调节导致的相变。A, B 在非运动细胞中,皮层张力和细胞间粘附强度之间的平衡可以导致密度无关的固体-液体转变。控制参数(形状指数 s0)决定了局部细胞重排的概率。在上皮细胞中,对应于正六边形的形状指数(3.72)标志着稳定性的丧失,并且刚性相变发生在对应于正五边形的形状指数(3.81[3])。C 细胞分裂可导致组织的流化,这是由于在有丝分裂细胞圆化过程中细胞间接触的丧失。在早期斑马鱼胚胎(受精后 4 小时)中,胚盘中心的细胞经历有丝分裂,显示细胞间接触长度减少和间质间隙增加,从而导致组织粘度和类流体行为暂时降低。然而,由于增强细胞间接触的非经典 Wnt 信号传导,边缘的细胞没有显示流化[8]。D 密集组织中的细胞损失(由于细胞死亡或分层)可导致细胞间连接的损失和组织粘度的降低,并导致类流体行为[46]。E 当在组织中被抑制时,由于肌动球蛋白收缩性(例如在八细胞阶段的早期小鼠胚胎中看到的)细胞的脉冲收缩,由于细胞间接触,导致长时间的力并增加整体机械完整性组织[50]。F 在临界值附近的粘附依赖性细胞连通性(例如斑马鱼胚层中的细胞连通性)的变化可导致刚性相变,如渗流理论所预测的。低于细胞连通性临界值的巨型刚性簇(红色)突然消失可导致组织粘度突然降低,使组织更像流体[56]。C-F 图中的垂直虚线(紫色)表示发生相变的临界点(在相关参数空间中)。
在某种程度上,作为固体标志的形状指数的存在,让人想起为晶体结构序列参数的定义。这表明尽管某种材料仍然是无序的,但可以通过观察其组成成分的形状来识别它是流体还是固体。人们可以认为这是依赖于模型的,确定状态的唯一相关方法是搞清楚机制(参见“原位检测相变的实验方法和挑战”部分和图 5)。
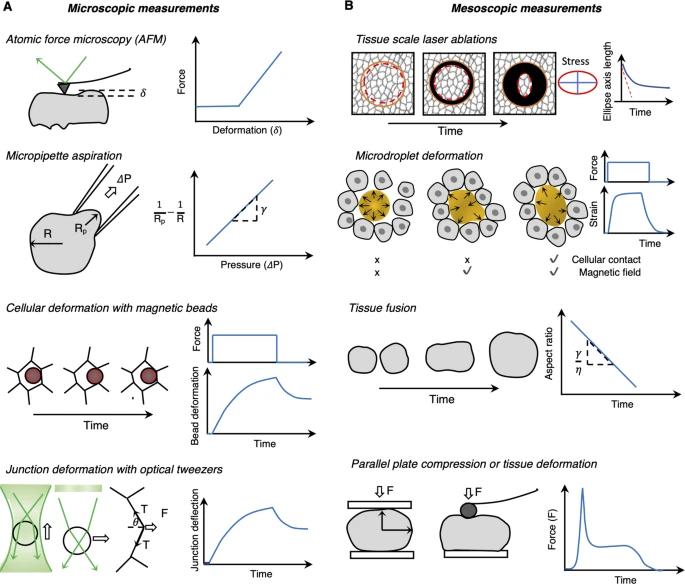
图 5:用于原位检测相变的流变测量(基于接触的方法)。A 一类显微技术旨在探测细胞尺度的流变学,其中测量通常需要机械装置或插入力探针并确定力-变形曲线。B 中观流变测量旨在测量超细胞特性,其依赖于利用力-变形曲线的相同原理,但在多细胞水平上诱导变形。
动力学相变
Vicsek 等人[31]首先从理论上探讨了自推进粒子系统中自发有序运动的出现。他们的工作受到生物系统的推动,模拟了一个以恒定的绝对速度运动的粒子系统,运动方向与粒子在给定半径附近的运动方向相同,但添加了一些随机扰动(图 3D)。通过进行数值模拟,他们观察到,这种简单的相互作用可以通过旋转对称的自发对称性破缺,导致从无转变到有限净转变的一种新相变——这被称为动力学相变(kinetic phase transition)。从概念上讲,这种相变可以看作类似于铁磁材料中的自旋排列,其中温度在运动方向上起到了随机扰动的相应作用,以实现速度排列。虽然这种转变不一定是固体-流体转变,但运动的对齐可以促进集体运动并促进凝固[32]。
一般来说,这种相变有助于理解细胞群内的集体细胞运动,具体取决于密度和噪声(即运动方向的扰动水平)(图 3C、D)。在密度和噪声很小的情况下,粒子可以形成群体,沿随机方向连贯运动;然而,如果密度高于一个临界阈值,则宏观上就会涌现出有序运动,其中所有粒子倾向于自发地沿同一方向运动(图 3C、D)。在金鱼角质细胞的集体迁移中已经观察到这种转变的生物学证据,其中成组移动的相互作用的细胞簇可以在临界密度附近形成[33]。
这种向有序状态的转变可以通过短程相互作用来解释,而无需任何关于邻居运动方向的明确信息。同样,当以高细胞密度铺板时,观察到由 MDCK 细胞组成的运动上皮中集体迁移细胞的速度场非常一致。最近在癌症方面的研究表明,当小 GTPase RAB5A 酶过表达时,细胞会表现出集体运动,让人联想到动力学(聚集性)相变。肿瘤球体中持续和协调的运动逐渐重塑了细胞外间质,进一步促进了癌细胞的集体侵袭和扩散。局部邻域内细胞运动方向的对齐很可能是趋向性或机械趋向性的结果,其表现为几种细胞直径的整体排列。
细胞接触/细胞波动的动态调制
温度是与相变相关的重要物理参数(框 1),因为它是对组成分子平均动能的量度。因此,与同一物质材料的液相或气相相比,固体中的分子移动较少,这实际上意味着凝固点的温度低于沸点。因此,温度被很好地作为固液转变的控制参数,特别是对于泡沫状系统从湿态到干态的固液“堵塞”。直到最近,才提出了一个被称为细胞抖动(cell jiggling)的等效参数来解释为有效温度[37,38]。与温度的运动学图像类似,细胞抖动是衡量细胞之间接触波动的指标,它是由多种分子过程引起的,这些分子过程可以调节细胞间粘附或细胞收缩性[39]。在本节中,我们提出了组织内的几个活跃过程,这些过程可能有助于细胞接触/细胞波动的变化,并将细胞抖动作为研究生物组织相变的独立控制参数(独立于细胞密度和应力)。
由于主动张力波动和堵塞相变引起的变化
多细胞生物中的固化可以看作是细胞运动和重新排列能力的丧失。这种过程强烈依赖于细胞能够产生的主动驱动波动。在斑马鱼中,细胞的逐渐凝固是轴伸长的基础。与雏鸡不同,这个过程不涉及增殖;相反,来自位于动物前部的中胚层祖区的间充质细胞在进入前胚体中胚层(后部)时逐渐失去重新排列的能力。这种流体到固体的转变类似于堵塞相变,当细胞进入前胚体中胚层时,它们会变成笼子(图 2B)。前胚体中胚层的细胞形成称为体节(somites)的结构,这些结构会产生动物的脊椎。为了绘制细胞尺度的机械特性,Mongera 等人[2]使用了可磁变形的油滴,这些油滴被注入斑马鱼胚胎的尾部。他们测量了使组织永久变形所需的应力量(屈服应力),并发现它在从后到前的方向上增加,表明后部更接近固体,而前部则逐渐接近固体。此外,控制固体-流体转变的张力波动幅度似乎可以调整,以使体节边界成形,体节内部是固体,而紧邻的组织是流体(图 2C)[40]。这些观察结果与考虑细胞外空间和细胞间接触处的主动张力的动态顶点模型一致。在张力波动的临界值上,该模型预测组织表现为流体。低于临界值,组织可以根据发育时间表现为流体或固体。如果形成结构的发育时间尺度大于(小于)应力弛豫时间尺度,则组织表现为流体(固体)[41]。在不同的情况下,通过胚胎干细胞的基于主体的模拟,也报告了在活性液滴的停滞聚结期间玻璃状或堵塞细胞行为[42, 43]。
细胞分裂和细胞死亡引起的变化
由于有丝分裂细胞的圆形化,细胞分裂通常会破坏细胞间的接触。组织特性可以根据组成细胞分裂的发生频率和同步性进行调节。与细胞分裂一起,细胞死亡可以引入通常是各向异性的应力源,因此能够产生有效的剪切粘度,导致组织表现为粘弹性流体,弛豫时间由分裂和细胞凋亡的速率设定[26]。上皮组织中的细胞分裂和细胞死亡都可能是导致组织局部流化的细胞重排的来源。在雏鸡胚胎中,细胞分裂在原肠胚形成过程中驱动细胞嵌入事件,如“由于主动驱动的细胞重排而导致的变化”一节中所讨论的。分裂细胞的连接重塑需要在其附近的低皮层肌动球蛋白,这样分裂细胞可以使临近细胞变形和位移。皮质的高转换导致低皮质刚性和低E-钙粘蛋白连接稳定性。F-肌动蛋白和肌球蛋白稳定性的增加会损害分裂介导的重排[25]。这种细胞水平的要求使组织更像液体,并使在雏鸡的原肠胚阶段中观察到大规模流动模式成为可能。
即使在非上皮组织的情况下,细胞分裂也可能是固液转变的驱动因素(图 4C)。例如,在合子基因组激活之前的大多数早期胚胎中,细胞分裂是同步的,并且取决于物种,细胞分裂的速率将决定组织特性变化的时间尺度。特别是对于斑马鱼胚胎,发育的前 25 小时几乎每 15 min 伴随着一次的同步细胞分裂,实际上意味着所有细胞每 15 min 经历一次有丝分裂细胞圆形化和松散的细胞间接触。这显然会影响组织及其相(固体状或液体状)的整体力学性能,具体取决于细胞是否相互接触。
随着发育的进行,细胞分裂可以成为组织特性区域化调节的来源,正如 Petridou 及其同事在斑马鱼原肠胚形成开始时胚层扩散的情况中观察到的那样[8]。他们表明,中央胚层分裂(分裂周期 12 和 13)期间的细胞圆化使组织更像液体。然而,边缘细胞在局部激活非经典 Wnt 信号(Wnt11-Fz7),从而增加细胞凝聚力以抵消有丝分裂对接触解体的影响。凝聚力或类固体行为是由 Wnt11-Fz7 信号依赖的肌动球蛋白收缩性增加引起的,这促进了 E-钙粘蛋白-肌动蛋白在细胞间接触边缘的定位[44]。作者进一步表明,这种空间受限的变化,是组织特性对于组织形态发生所至关重要的,因为在wnt11/slb 突变体胚胎中看到的类流体行为的均匀变化,会导致胚层变薄[45]。
在生长的组织中,细胞拥挤密集,这不可避免地导致密集堆积和潜在的凝固。在上皮组织中,细胞分层(细胞死亡之前的过程)被证明可以抵消过度拥挤[46]。在细胞邻居将它们挤出之前,细胞因细胞间连接的丧失和顶端面积的减少而分层(图 4D)。正如先前理论所建议的[47,48],拥挤可以机械地反馈到细胞上,以缓冲组织生长并确保组织稳态。例如,一种被称为接触抑制的细胞增殖抑制形式,已被证明是由密度变化引起的。拥挤和生长之间的机械反馈受损可能有利于增生和肿瘤形成,这通常伴随着组织固态化的过程。
皮层脉冲和波引起的变化
在早期小鼠胚胎中,当细胞脱离粘附接触时,细胞的脉冲收缩在短时间内表现为周期性皮层波。然而,在组织中,这种细胞自主脉冲收缩,当受到限制或抑制时,由于钙粘蛋白介导的细胞间接触会被重新定向远离细胞连接,从而在长时间尺度上产生组织水平的力。特别是在早期(八细胞阶段)小鼠胚胎的情况下,这种肌动球蛋白收缩性会导致压实,从而增加胚胎的整体机械完整性[50]。肌动蛋白-肌球蛋白网络的这种脉冲收缩已被证明可驱动果蝇中的细胞形状变化[51,52,53,54],并由于细胞和组织的粘弹性导致整体组织流动,使得它们尽管存在瞬态力但仍可永久变形。总体而言,推测细胞粘附和细胞自主脉动之间的平衡可以导致多细胞系统中的长期力是合理的,这些力可以通过动态调节细胞接触/细胞来影响组织的类固体或类流体行为。
由于细胞间连通性、渗流和网络相变引起的变化
网络中链接连通性的变化会导致刚性的宏观变化。连通性的程度类似于液体通过多孔物质渗透的难易程度:更高的连通性意味着没有液体不能自由通过。这种情况被称为通过网络的“刚性渗流”(percolation of rigidity,区别于液体)。与物理学中这种经过充分研究的过程类似,细胞间连通性的变化可以改变组织刚性,即可能导致渗流。在无限网络中,渗流理论预测刚度在临界链接密度之上的相变(框 2 和[55])。对于组织等有限尺寸的多细胞系统,当细胞连接性达到阈值时,当应力在系统尺寸上传播时,粘度是否会发生突然变化并没有显示出来。然而,最近在斑马鱼胚盘中的一项研究证明了一种真正的刚性相变,这是由粘附介导的细胞连接性变化引起的(图 4F 和[56])。作者表明,当每个细胞的平均接触量较低时,不出所料,组织表现为流体但高于连接性巨大刚性簇(框 2)的临界阈值,并跨越整个网络以抵抗变形,从而使组织呈现行为更像固体。通过组合遗传扰动来改变细胞连通性(通过降低 E-cadherin 表达水平或通过下调 Wnt/PCP 途径(图 4C)或通过改变细胞命运规范),表明细胞连通性是一个可靠的参数使用刚性渗流分析预测组织粘度的变化。这种网络相变是生物系统在形态发生过程中如何利用相变来雕刻组织的另一个例子[57]。相反,接近临界会导致不稳定性,这在生物系统中通常不会观察到,我们设想未来的研究将解决这一难题,为我们理解相变在生物系统中的作用开辟新途径。
框2
对称性和对称性破缺(Symmetry and symmetry breaking)。物体的对称性(几何学)是指物体在几何变换下的不变性。例如,如果一个对象在旋转之后可以映射到它自己,那么它就是旋转对称的。对称性破缺是一个过程,通常由微小的波动引起,使系统从一个对称的状态变成一个不那么对称的状态。最终状态不同于初始状态,需要更多的特征来描述它。例如,液态水在各个方向上看起来都是一样的。当它变成一片雪花时,它失去了旋转对称,只在六个方向上看起来一样。
玻璃相变和堵塞相变(Glass transition and jamming transition)。玻璃转变是指从粘性液体到非晶态(无序的,非晶态)固体的转变。堵塞相变是在不使用温度调节下的玻璃相变,用来表示从液态或“松软”状态到刚性状态的转变。它涵盖了不同的情况,如交通堵塞的形成,或沙子/糖果的堆积,或玻璃相变。伴随而来的是各组分活力的急剧下降。因此可以认为它是一种动力学相变。堵塞相变是体现在系统“冻结”中的一种普遍现象。例如,当一定体积的沙子倒在桌子上,流动会自发停止,并得到一个机械稳定的沙堆。当胶体体积分数较大时,胶体组成成分表现出相同类型的转变:在高浓度的硬颗粒体系中,颗粒被束缚在由其邻近颗粒形成的瞬态“笼子”中。在接近堵塞的情况下,正式“笼子”的重新排列控制(减缓)了动力学[105]和变形抗力。运动速度的减慢与粒子集群有关,其特征尺寸随拥挤程度的增加而增大[106]。干扰系统的一个显著特性是控制参数的适度变化(例如密度的增加)导致有效粘度的发散和有限刚度的出现。有关堵塞及其来源在生物学背景下的详细回顾,请参阅参考文献[20]。
渗流和网络相变(Percolation and network phase transitions)。在网络中,节点及链路的增加或删除可以改变网络的宏观行为。在临界链路密度以下,由于网络内连续变形的数量增加,网络可以分解成较小的连接簇。或者,当超过这个临界链路密度,网络将表现为刚性,换句话说,刚性渗流到系统中[55,107,108,109,110]。网络结构的这种变化导致了相变:在临界密度以下,网络将具有大的松软区域和一些刚性节点团簇,而在这个临界值以上,则能够跨越整个网络的刚性节点团簇会突然涌现,称为巨大刚性团簇(Giant Rigid Cluster)[56,110,111]。渗流理论预测这个临界值是最大平均接触数的2/3。
三、原位检测相变的实验方法和挑战
惰性材料中的固体到流体转变伴随着组成粒子的长程序列的突然变化。然而,如前所述,生物材料在很大程度上仍然是无序的。这对识别它是否表现为流体或固体特征提出了挑战,因为没有明显的特性可以最终确定材料的相,并因此判断是否发生相变。然而,相变的一个显着特征是材料性质的剧烈变化。例如,堵塞相变伴随着粘度的发散和有限刚度的获得。因此,在生物组织或细胞群中检测此类相变的一种明显方法是对其进行机械探测并在原位测量其流变特性。此类测量可以称为流变测量(rheological measurements)。
另一种通过关注动力学来推断组织相的方法,即成像并跟踪细胞行为和细胞形状。组成细胞的形状可以提供组织机械状态的线索。对单个细胞或细胞群的轨迹及其扩散特性的分析可以提供生物组织相特性的信息。例如,流体特点是颗粒自由扩散并能够在剪切作用下相互通过,而玻璃(堵塞的固体)显示出限制颗粒扩散的“笼子效应”。这种类型的测量可以称为运动学测量。
流变测量(基于接触的方法)
流变实验是材料科学中的经典实验,包括测量材料在外力作用下的流动和变形。它们在生物系统中的实施通常具有挑战性,但近年来已经能看到既有技术的适应性以及新技术的开发(详细综述参见[58])。这些类型的测量通常需要一个机械装置或插入力探针和力-变形曲线的确认(图 5)。根据测量的尺度,我们可以将它们分类为微观(在细胞尺度上测量流变)或中观测量(在细胞外或组织尺度上)。
微观测量类别包括原子力显微镜(原子力显微镜)[10]、微量移液器抽吸[59]、光镊[60]、亚细胞激光消融后的细胞顶点位移[52]和磁珠等方法,所有这些都依赖于基于力-位移(变形)曲线的推断(图 5A)。另一种新兴的方法是布里渊显微镜[61],它是一种测量组织粘弹性特性的无标记和无接触方法。使用其中一些技术的经典示例(详见[8])包括单细胞力谱(SCFS)[62]测量斑马鱼胚胎中胚层祖细胞的粘附,融合实验以推断组织表面张力差异[63],原子力显微镜测量非洲爪蟾头部中胚层的弹性[64]或内皮、心肌和骨骼肌的粘弹性[65],光镊在早期果蝇上皮中变形细胞连接和粘性耗散[9,66],布里渊显微镜来绘制正常和患病的人类角膜弹性特征[67]或斑马鱼脊索中细胞外基质刚度[68]和微量移液器抽吸以测量不同脊椎动物胚胎中的组织粘弹性如斑马鱼囊胚[8]、非洲爪蟾[69]、鸡[70]和小鼠[50]。
中观流变测量旨在测量细胞外或多细胞的特性,这些特性与微观测量一样依赖于力-变形曲线,但在多细胞水平上诱导变形(图 5B)。组织级激光消融可以探测伤口闭合的各向异性[71]。平行板压缩技术借鉴了材料科学中使用的经典压缩技术,但在微观尺度上,表面张力效应主导组织变形和形状[72]。微滴变形可以测量刚度各向异性[73],并且可以使用均匀磁场下的铁磁流体液滴研究组织对大变形的机械响应[74]。例如,Mongera 等人[2]在斑马鱼胚胎的尾部注射了这种可磁变形的油滴,并测量了使组织永久变形所需的应力(屈服应力)沿着身体轴线的从后往前增加,表明在前部更呈流体状,后部逐渐呈固体状。藻酸盐或聚丙烯酰胺中的软弹性微球可用于测量各向同性(拉伸和压缩)和剪切应力[75,76]。原子力显微镜等技术也已被用于中观测量,方法是将大珠连接到原子力显微镜的悬臂以测量生物组织尺度的变形[64]。
在这两类测量中,为了检测相变的证据,因此至关重要的是,尽可能在自然状态下扰动和探测组织,并且最好是推断组织的动态状态而非被动测量其组成细胞的机械特性。然而,这并不总是可行的,因此需要为给定系统谨慎选择技术和分析方法,因为推断的特性可能会取决于所使用力探针的大小和速率、所用探头的几何形状和配置以及所探测的组织区域[77]。
运动学测量(非接触式方法)
此类测量依赖于对自然环境中细胞的运动和形状的分析,以对组织的机械状态深入了解(图 6)。实时成像、荧光或其他方式是这类测量的关键工具。
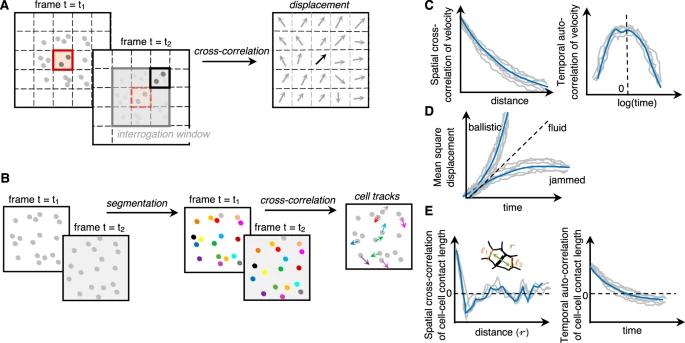
图 6:用于原位检测相变的运动学测量(非接触方法)。A、B 速度测量可以通过粒子图像测速或a.基于粒子集合的运动(全局流动模式)b.通过分割和跟踪的单个粒子的运动等相关技术来完成。C 诸如空间互相关和时间自相关等速度的高阶测量允许分别推断集体细胞运动的弛豫时间和相关长度,这两者都可用于检测细胞单层中的相(堵塞)转变[4,78]。D 均方根位移可以从粒子(细胞)的各个轨道推断出来,并可用于推断“笼式”与“非笼式”动力学。E 可以对分割图像中的细胞间接触长度 (l) 进行高阶测量:间隔 r 的细胞间接触长度的空间互相关或细胞间接触长度的时间自相关可用于检测相(堵塞)转变的“抖动”特征量[2]。
例如,速度测量和高阶测量,如速度空间相关性和速度时间相关性,可用于检测相变。速度自相关提供了弛豫时间[78],预计弛豫时间会在接近玻璃化转变时显着增加。速度空间相关性能够捕捉集体运动及其相关长度。这些测量已用于表征细胞单层中的集体运动和堵塞相变。我们预计它们将有效地应用于体内和三维系统,因为该方法是非侵入性的并且依赖于成像。
四、超越相变:组织刚度的缓慢变化
生物组织的材料特性的变化可以在几个小时到几天的时间尺度上逐渐发生,而不存在任何像典型相变一样的临界点。尽管速度缓慢,但力学性能的变化可能是巨大的。在这种情况下,组织的凝固或流化可能是由细胞内成分和细胞外环境的变化造成的。
细胞组成引起的变化
细胞中水含量是决定细胞力学性能的关键因素。特别是对于具有细胞壁的植物细胞,由于水的流入/流出,细胞内的静水压力可高达 20 个大气压(约2 MPa[80])。这种压力,称为膨胀压力,因此对于驱动局部形态发生和组织的整体完整性和刚度至关重要。与细胞活动的动物组织不同,植物组织通过调节细胞壁的材料特性和膨胀压力来控制其硬度,从而导致其结构的大规模变形。虽然由于细胞组成的这种变化而导致突发相变的证据尚不清楚,但调节水量仍然是控制整体组织刚度的有力机制。例如,花发育过程的花药开放释放成熟花粉(该过程称为花药开裂[81,82]),或由于花瓣扩张而使花蕾开放,也暴露其心皮上的柱头进行授粉[83],两者都是由特定细胞群的水合和脱水序列引起的,这些细胞群在空间上调节相对刚度,从而导致受控的组织整体变形(图7A、B)。在分子水平上,水的流入和流出受K+/Na+离子转运蛋白和水通道蛋白分布的空间调控控制。最近有报道称,调节水渗透是在受限微环境中控制细胞迁移的一种机制[84]。Stroka 等人的工作表明受限的肿瘤细胞在细胞膜中建立了 Na+/H+ 泵和水通道蛋白的极化分布,从而在细胞的前缘和后缘之间产生水和离子流梯度,从而实现细胞排水。
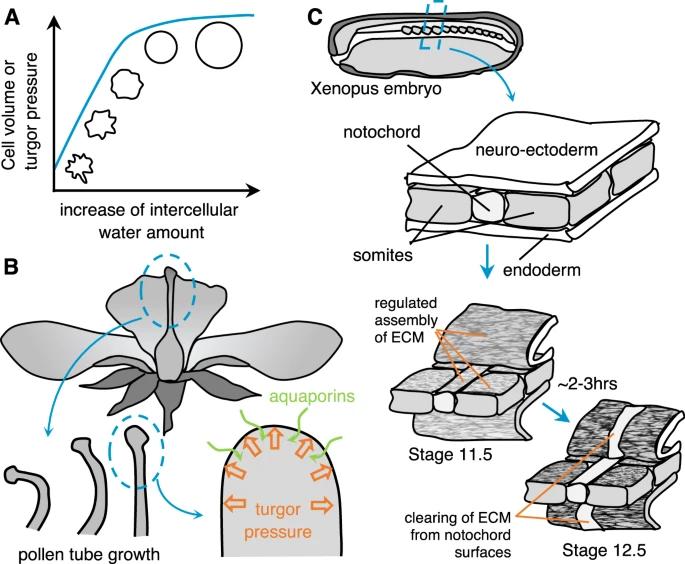
图 7:细胞组成和细胞外环境变化导致的组织刚度变化。A,B 植物细胞中存在的水量,建立膨胀压力(A),并驱动组织的局部形态发生和整体完整性和刚性,例如花粉管在花中的生长(B)[80]。C 细胞外基质成分,例如纤连蛋白原纤维,划定了组织边界,例如发育中的非洲爪蟾胚胎中的神经/中胚层界面之间[87]。
细胞外环境引起的变化
细胞在群体水平上的类固体或类液体行为也可以由细胞外在因素决定。在这方面,细胞外基质 (ECM) 是细胞分泌的一个关键决定因素,它通过对细胞行为的机械化学控制影响集体细胞动力学。整体组织刚度的变化,可能是分泌细胞外基质数量和方向的直接结果。骨和软骨组织是典型的例子,其间充质细胞的软聚集体分化成大型肥大软骨细胞,合成 X 型胶原蛋白并钙化它们周围的细胞外基质[85]。随后,这些软骨细胞的协调凋亡以及它们被进一步分泌 I 型胶原蛋白的成骨细胞替代,改变了组织的整体刚度进而形成刚性骨结构[85]。在骨关节炎等疾病中,成骨细胞不能分泌足够的细胞外基质,从而影响骨骼的力学性能。另一个显著例子是分泌的几丁质基质,它与碳酸钙结合,成为一种固体材料,在节肢动物如甲壳类动物和软体动物的外壳中构成外骨骼[86]。
骨骼、软骨和外骨骼等固体结构的形成是渐进的,并持续数天。基质沉积在较小时间尺度上的效应及其影响已在癌症等疾病的发展和疾病中得到广泛研究。在非洲爪蟾胚胎中观察到了这种发育影响的第一个证据。身体轴线沿 A/P 轴的延伸伴随着背侧退化边缘的硬化。沿 A/P 轴方向的硬化程度大于沿中外侧轴方向。研究者提出组织硬化是由细胞骨架的增强或细胞外基质的沉积引起的,这将有助于维持刚性轴[88]。有证据表明纤连蛋白原纤维在组织边界处形成(图 7C)[87]。它们的模式(pattern)可以控制力学性能,如弹性模量,但也可以其润滑作用以减少形态发生过程中组织之间的剪切。
更一般地说,细胞外环境的变化会影响组织的分化、方向和运动,所有这些都可能导致组织硬化[89]。例如,最近表明,在果蝇卵泡中,通过 Src 酪氨酸激酶感知的基底膜刚度,改变了卵泡前部细胞连接的 E-钙粘蛋白运输[90]。因此,在平均细胞形状或定向细胞分裂没有任何变化的情况下,细胞在无边管状上皮中重新定向,这主要是由于细胞外基质的定向,从而控制了三维组织的形状和刚度。细胞外基质控制细胞运动的另一种情况是先前讨论的鸡胚胎前胚体中胚层 (PSM) 中细胞分级运动的情况。在此,间充质细胞在单个细胞水平上的类气体行为(导致无方向布朗运动),会由于底层细胞外基质的运动,而转化为类似于液体的分级流动模式。
已经有大量关于细胞外基质成分和生物物理特性如何诱导上皮-间质转化(EMT)和促进迁移行为的文献 [91,92]。肿瘤生长和扩散是否类似于相变,是十年来一个持续争议的问题(例如参考文献[93])。发育过程和癌症之间存在惊人的相似之处,它们对细胞外基质刚度的敏感性,以及更普遍地对环境刚度的敏感性。细胞外基质刚度可能会沿着细胞外基质刚度梯度促进肿瘤生长、扩散和转移,这一过程称为硬轴定位(durotaxis)[94]:同样,底物刚度可以通过在体内神经嵴形成过程中促进上皮-间质转化来触发集体细胞迁移。在后一种情况下,神经嵴细胞迁移是对支撑中胚层硬化的反应。因此,组织类液体或类固体行为受内在和外在因素的调节。
通常,像这样的凝固实例在几天的时间尺度上逐渐发生,没有任何典型的相变临界过度点。
在形态发生过程中,组织受到周围组织和环境(细胞外基质、壳、卵周膜)的外力和机械约束。几个团队已经开始解决组织如何机械地响应这些力和约束的问题。最近的报告表明,力学性能,例如硬化是由外部条件引起的。拉伸引起的硬化已被提出作为一种限制变形后形状变化的机制。使用拉伸装置,Duda 等人[95]对果蝇翼盘上皮施加外部载荷并观察组织的机械响应,沿拉伸方向形成极化的肌动球蛋白索。肌动球蛋白网络通过采用不同的配置来感知和响应几何与机械约束[96]。它们可以形成环形或星形,这反过来又可以引导力的产生及力学性能改变。在改变边界的不同位置进行激光切口或烧灼是确定外部组织对力学性能的影响的一种方法[52,96,97]。外部条件的调制是否会导致相变还有待证明。应变硬化可以被视为一种减少甚至防止组织变形的一般凝固机制。
五、生物与非生物系统类比的局限及未来方向
在生命系统中的集体效应与物理惰性系统中众所周知的相变之间建立类比具有说服力。例如,将经典的三轴堵塞相图应用于细胞单层的情况,有助于突出可能控制组织中流固转变的关键参数,即细胞运动/细胞抖动,细胞间粘附性的倒数和细胞密度,作为温度、应力和体积分数或密度的对应物[37]。然而,参数的唯一性是值得怀疑的。在体积分数接近[1]的汇合单层中,该参数与描述堵塞相变无关,而是提出了以细胞形状为特征的参数[17]。这说明了搜索控制参数并测试它们相关性的必要性。
同样,虽然很明显,波动通常会促进组织的流化,但探索这些波动(由热造成的或非由热造成的)的确切性质至关重要。在细胞运动和(有效)温度之间进行类比可能会产生误导,因为由主动驱动的波动(导致细胞运动)和热波动在本质上是非常不同的[99,100,101]。与热力学系统中普遍存在的玻尔兹曼分布所给出的稳态相比,主动驱动的应力导致在质量上不同的稳态。这阻碍了生命系统中“有效温度”的定义[102]。分子马达产生的主动波动决定了形状波动的特征时间,进而导致细胞重排和细胞运动。我们对分子和细胞尺度之间的关系知之甚少,因此开发能将它们联系起来的物理模型和理论非常重要,这将有助于我们定义控制相变的有效参数。相变概念在生物系统中的应用受到生物非平衡性质的限制。观察到的生命系统状态都不是平衡状态,而是从非平衡动力学中涌现的瞬态或稳态。在胚胎形态发生过程中,由基因表达、动态粘附和细胞骨架结构的变化引起的一系列状态可能会使相变的识别存在风险甚至误导。尽管如此,描述这些状态的材料特性仍然必不可少,我们相信在某些情况下它们将揭示行为层面的转变,其中一些将是相变的标志。转变的位置将取决于生物系统的具体情况,但是,正如其他动态系统[103]所显示的那样,很可能会出现对细节不敏感的特征。
一项吸引人但令人生畏的任务是将控制相变的物理参数与生物实体联系起来。尝试将蛋白质功能与物理参数匹配存在固有风险,因为许多蛋白质有助于多种物理特性。例如,肌球蛋白-II 的活性增强细胞波动,但也能够作为肌动蛋白交联剂改变细胞刚度。牢记这一点,使用急性遗传和分子扰动结合的机械测量与成像技术,对于破译重要参与者、确定系统在相图中的位置至关重要,并有望探索参数空间。
生物系统在力学性能变化的时间尺度和变形的时间尺度之间表现出强烈的耦合,因此与惰性材料相比,应力在相变中可以发挥非常不同的作用,尤其是堵塞相变。反馈在从分子到组织尺度的机械和生化信号之间发挥作用。此外,大小和几何形状可以对多细胞系统的集体行为发挥关键作用。这使得生命系统相比于物理惰性系统显得非常特别。
未来的工作应该深入了解几何、力学和生物化学的多尺度耦合,以及它们如何整合以定义多细胞系统的集体行为,以及控制生物体形成过程中相变的参数。
参考文献
1.Saw, T. B. et al. Topological defects in epithelia govern cell death and extrusion. Nature 544, 212–216 (2017).
2.Mongera, A. et al. A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature 561, 401–405 (2018).
3.Bi, D., Lopez, J. H., Schwarz, J. M. & Manning, M. L. A density-independent rigidity transition in biological tissues. Nat. Phys. 11, 1074–1079 (2015).
4.Garcia, S. et al. Physics of active jamming during collective cellular motion in a monolayer. Proc. Natl Acad. Sci. USA 112, 15314–15319 (2015).
5.Iyer, K., Piscitello-Gómez, R., Paijmans, J., Jülicher, F. & Eaton, S. Epithelial viscoelasticity is regulated by mechanosensitive E-cadherin turnover. Curr. Biol. 29, 578–591 (2019).
6.Malet-Engra, G. et al. Collective cell motility promotes chemotactic prowess and resistance to chemorepulsion. Curr. Biol. 25, 242–250 (2015).
7.Cheung, K. J. & Ewald, A. J. A collective route to metastasis: seeding by tumor cell clusters. Science 352, 167–169 (2016).
8.Petridou, N. I. & Heisenberg, C. Tissue rheology in embryonic organization.
EMBO J. 38, e102497 (2019).
9.Clément, R., Dehapiot, B., Collinet, C., Lecuit, T. & Lenne, P. F. Viscoelastic dissipation stabilizes cell shape changes during tissue morphogenesis. Curr. Biol. 27, 3132–3142 (2017).
10.Krieg, M. et al. Tensile forces govern germ-layer organization in zebrafish.
Nat. Cell Biol. 10, 429–436 (2008).
11.Angelini, T. E. et al. Glass-like dynamics of collective cell migration. Proc. Natl Acad. Sci. USA 108, 4714–4719 (2011).
12.Bi, D., Yang, X., Marchetti, M. C. & Manning, M. L. Motility-driven glass and jamming transitions in biological tissues. Phys. Rev. X 6, 021011 (2016).
13.Brookes, N. H. Riding the cell jamming boundary: geometry, topology, and phase of human corneal endothelium. Exp. Eye Res. 172, 171–180 (2018).
14.Chepizhko, O. et al. From jamming to collective cell migration through a boundary induced transition. Soft Matter 14, 3774–3782 (2018).
15.Delarue, M. et al. Self-driven jamming in growing microbial populations. Nat. Phys. 12, 762–766 (2016).
16.Park, J. A. et al. Unjamming and cell shape in the asthmatic airway epithelium.
Nat. Mater. 14, 1040–1048 (2015).
17.Park, J. A., Atia, L., Mitchel, J. A., Fredberg, J. J. & Butler, J. P. Collective migration and cell jamming in asthma, cancer and development. J. Cell Sci. 129, 3375–3383 (2016).
18.Sadati, M., Taheri Qazvini, N., Krishnan, R., Park, C. Y. & Fredberg, J. J. Collective migration and cell jamming. Differentiation 86, 121–125 (2013).
19.Schötz, E. M., Lanio, M., Talbot, J. A. & Manning, M. L. Glassy dynamics in three-dimensional embryonic tissues. J. Royal Soc. Interf. 10, 20130726 (2013).
20.Lawson-Keister, E. & Manning, M. L. Jamming and arrest of cell motion in biological tissues. Current Opinion in Cell Biology 72, 146–155 (2021).
21.Bénazéraf, B. et al. A random cell motility gradient downstream of FGF controls elongation of an amniote embryo. Nature 466, 248–252 (2010).
22.Bertet, C., Sulak, L. & Lecuit, T. Myosin-dependent junction remodelling controls planar cell intercalation and axis elongation. Nature 429, 667–671 (2004).
23.Krajnc, M., Dasgupta, S., Ziherl, Pcv & Prost, J. Fluidization of epithelial sheets by active cell rearrangements. Phys. Rev. E 98, 022409 (2018).
24.Blankenship, J. T., Backovic, S. T., Sanny, J. S. S., Weitz, O. & Zallen, J. A. Multicellular rosette formation links planar cell polarity to tissue morphogenesis. Dev. Cell 11, 459–470 (2006).
25.Firmino, J., Rocancourt, D., Saadaoui, M., Moreau, C. & Gros, J. Cell division drives epithelial cell rearrangements during gastrulation in chick. Dev. Cell 36, 249–261 (2016).
26.Ranft, J. et al. Fluidization of tissues by cell division and apoptosis. Proc. Natl Acad. Sci. USA 107, 20863–20868 (2010).
27.Sato, K. et al. Left-right asymmetric cell intercalation drives directional collective cell movement in epithelial morphogenesis. Nat. Commun. 6, 1–11 (2015).
28.Cohen-Addad, S., Höhler, R. & Pitois, O. Flow in foams and flowing foams.
Annu. Rev. Fluid Mech. 45, 241–267 (2013).
29.Farhadifar, R., Röper, J., Aigouy, B., Eaton, S. & Jülicher, F. The influence of cell mechanics, cell-cell interactions, and proliferation on epithelial packing. Curr. Biol. 17, 2095–2104 (2007).
30.Wang, X. et al. Anisotropy links cell shapes to tissue flow during convergent extension. Proc. Natl Acad. Sci. USA https://www.pnas.org/content/early/ 2020/05/27/1916418117.full.pdf (2020).
31.Vicsek, T., Czirk, A., Ben-Jacob, E., Cohen, I. & Shochet, O. Novel type of phase transition in a system of self-driven particles. Phys. Rev. Lett. 75, 1226–1229 (1995).
32.Giavazzi, F. et al. Flocking transitions in confluent tissues. Soft Matter 14, 3471–3477 (2018).
33.Szabó, B. et al. Phase transition in the collective migration of tissue cells: experiment and model. Phys. Rev. E 74, 061908 (2006).
34.Petitjean, L. et al. Velocity fields in a collectively migrating epithelium.
Biophys. J. 98, 1790–1800 (2010).
35.Palamidessi, A. et al. Unjamming overcomes kinetic and proliferation arrest in terminally differentiated cells and promotes collective motility of carcinoma. Nat. Mater. 18, 1252–1263 (2019).
36.Lecaudey, V. & Gilmour, D. Organizing moving groups during morphogenesis. Curr. Opin. Cell Biol. 18, 102–107 (2006).
37.Sadati, M., Taheri Qazvini, N., Krishnan, R., Park, C. Y. & Fredberg, J. J. Collective migration and cell jamming. Differentiation 86, 121–125 (2013).
38.Liu, A. & Nagel, S. Jamming is not just cool any more. Nature 396, 21–22 (1998).
39.Fodor, É. et al. Spatial fluctuations at vertices of epithelial layers: quantification of regulation by Rho pathway. Biophys. J. 114, 939–946 (2018).
40.Shelton, E. R. et al. Stress-driven tissue fluidization physically segments vertebrate somites. Preprint at https://www.biorxiv.org/content/10.1101/ 2021.03.27.437325v1 (2021).
41.Kim, S., Pochitaloff, M., Stooke-Vaughan, G. A. & Campàs, O. Embryonic tissues as active foams. Nat. Phys. https://doi.org/10.1038/s41567-021-01215-1 (2021).
42.Oriola, D. et al. Arrested coalescence of multicellular aggregates. Preprint at https://arxiv.org/abs/2012.01455 (2020).
43.Ongenae, S., Cuvelier, M., Vangheel, J., Ramon, H. & Smeets, B. Activity- induced fluidization of arrested coalescence in fusion of cellular aggregates. Preprint at https://www.biorxiv.org/content/early/2021/02/26/ 2021.02.26.433001.full.pdf (2021).
44.Maître, J. L. et al. Adhesion functions in cell sorting by mechanically coupling the cortices of adhering cells. Science 338, 253–256 (2012).
45.Heisenberg, C. P. et al. Silberblick/Wnt11 mediates convergent extension movements during zebrafish gastrulation. Nature 405, 76–81 (2000).
46.Marinari, E. et al. Live-cell delamination counterbalances epithelial growth to limit tissue overcrowding. Nature 484, 542–545 (2012).
47.Shraiman, B. I. Mechanical feedback as a possible regulator of tissue growth.
Proc. Natl Acad. Sci. USA 102, 3318–3323 (2005).
48.Hufnagel, L., Teleman, A. A., Rouault, H., Cohen, S. M. & Shraiman, B. I. On the mechanism of wing size determination in fly development. Proc. Natl Acad. Sci. USA 104, 3835–3840 (2007).
49.Puliafito, A. et al. Collective and single cell behavior in epithelial contact inhibition. Proc. Natl Acad. Sci. USA 109, 739–744 (2012).
50.Maître, J. L., Niwayama, R., Turlier, H., Nedelec, F. & Hiiragi, T. Pulsatile cell- autonomous contractility drives compaction in the mouse embryo. Nat. Cell Biol. 17, 849–855 (2015)..
51.Martin, A. C., Kaschube, M. & Wieschaus, E. F. Pulsed contractions of an actin-myosin network drive apical constriction. Nature 457, 495–499 (2009).
52.Rauzi, M. et al. Embryo-scale tissue mechanics during Drosophila gastrulation movements. Nat. Commun. 6, 1–12 (2015).
53.Roh-Johnson, M. et al. Triggering a cell shape change by exploiting preexisting actomyosin contractions. Science 335, 1232–1235 (2012).
54.Solon, J., Kaya-Çopur, A., Colombelli, J. & Brunner, D. Pulsed forces timed by a ratchet-like mechanism drive directed tissue movement during dorsal closure. Cell 137, 1331–1342 (2009).
55.Moukarzel, C. & Duxbury, P. M. Comparison of rigidity and connectivity percolation in two dimensions. Phys. Rev. E 59, 2614–2622 (1999).
56.Petridou, N. I., Corominas-Murtra, B., Heisenberg, C.-P. & Hannezo, E. Rigidity percolation uncovers a structural basis for embryonic tissue phase transitions. Cell 184, 1914–1928 (2021).
57.Krotov, D., Dubuis, J. O., Gregor, T. & Bialek, W. Morphogenesis at criticality.
Proc. Natl Acad. Sci. USA 111, 3683–3688 (2014).
58.Sugimura, K., Lenne, P. F. & Graner, F. Measuring forces and stresses in situ in living tissues. Development 143, 186–196 (2016).
59.Guevorkian, K., Colbert, M. J., Durth, M., Dufour, S. & Brochard-Wyart, F. Aspiration of biological viscoelastic drops. Phys. Rev. Lett. 104, 218101 (2010).
60.Zhang, H. & Liu, K. K. Optical tweezers for single cells. J. Royal Soc. Interf. 5, 671–690 (2008).
61.Prevedel, R., Diz-Muñoz, A., Ruocco, G. & Antonacci, G. Brillouin microscopy: an emerging tool for mechanobiology. Nat. Methods 16, 969–977 (2019).
62.Benoit, M., Gabriel, D., Gerisch, G. & Gaub, H. E. Discrete interactions in cell adhesion measured by single-molecule force spectroscopy. Nat. Cell Biol. 2, 313–317 (2000).
63.Schötz, E.-M. et al. Quantitative differences in tissue surface tension influence zebrafish germ layer positioning. HFSP J. 2, 42–56 (2008).
64.Barriga, E. H., Franze, K., Charras, G. & Mayor, R. Tissue stiffening coordinates morphogenesis by triggering collective cell migration in vivo. Nature 554, 523–527 (2018).
65.Mathur, A. B., Collinsworth, A. M., Reichert, W. M., Kraus, W. E. & Truskey,
G. A. Endothelial, cardiac muscle and skeletal muscle exhibit different viscous and elastic properties as determined by atomic force microscopy. J. Biomech. 34, 1545–1553 (2001).
66.Bambardekar, K., Clement, R., Blanc, O., Chardes, C. & Lenne, P.-F. Direct laser manipulation reveals the mechanics of cell contacts in vivo. Proc. Natl Acad. Sci. USA 112, 1416–1421 (2015).
67.Yun, S. H. & Chernyak, D. Brillouin microscopy: assessing ocular tissue biomechanics. Curr. Opin. Ophthalmol. 29, 299 (2018).
68.Bevilacqua, C., Sánchez-Iranzo, H., Richter, D., Diz-Muñoz, A. & Prevedel, R. Imaging mechanical properties of sub-micron ECM in live zebrafish using Brillouin microscopy. Biomed. Opt. Express 10, 1420 (2019).
69.von Dassow, M. & Davidson, L. A. Natural variation in embryo mechanics: gastrulation in Xenopus laevis is highly robust to variation in tissue stiffness. Dev. Dyn. 238, 2–18 (2009).
70.Majkut, S. et al. Heart-specific stiffening in early embryos parallels matrix and myosin expression to optimize beating. Curr. Biol. 23, 2434–2439 (2013).
71. Bonnet, I. et al. Mechanical state, material properties and continuous description of an epithelial tissue. J. R. Soc. Interface 9, 2614–2623 (2012).
72.Foty, R., Pfleger, C., Forgacs, G. & Steinberg, M. Surface tensions of embryonic tissues predict their mutual envelopment behavior. Development 122, 1611–1620 (1996).
73.Campàs, O. et al. Quantifying cell-generated mechanical forces within living embryonic tissues. Nat. Methods 11, 183–189 (2014).
74.Serwane, F. et al. In vivo quantification of spatially varying mechanical properties in developing tissues. Nat. Methods 14, 181–186 (2017).
75.Mohagheghian, E. et al. Quantifying compressive forces between living cell layers and within tissues using elastic round microgels. Nat. Commun. 9, 1878 (2018).
76.Dolega, M. E. et al. Cell-like pressure sensors reveal increase of mechanical stress towards the core of multicellular spheroids under compression. Nat. Commun. 8, 14056 (2017).
77.Wu, P. H. et al. A comparison of methods to assess cell mechanical properties.
Nat. Methods 15, 491–498 (2018).
78.Nnetu, K. D., Knorr, M., Pawlizak, S., Fuhs, T. & Käs, J. A. Slow and anomalous dynamics of an MCF-10A epithelial cell monolayer. Soft Matter 9, 9335–9341 (2013).
79.Angelini, T. E. et al. Glass-like dynamics of collective cell migration. Proc. Natl Acad. Sci. USA 108, 4714–4719 (2011).
80.Beauzamy, L., Nakayama, N. & Boudaoud, A. Flowers under pressure: ins and outs of turgor regulation in development. Ann. Bot. 114, 1517–1533 (2014).
81.Scott, R. J., Spielman, M. & Dickinson, H. G. Stamen structure and function.
Plant Cell 16, S46–S60 (2004).
82.Nelson, M. R. et al. A biomechanical model of anther opening reveals the roles of dehydration and secondary thickening. N. Phytologist 196, 1030–1037 (2012).
83.van Doorn, W. G. & van Meeteren, U. Flower opening and closure: a review. J. Exp. Bot. 54, 1801–1812 (2003).
84.Stroka, K. M. et al. Water permeation drives tumor cell migration in confined microenvironments. Cell 157, 611–623 (2014).
85.Egawa, S., Miura, S., Yokoyama, H., Endo, T. & Tamura, K. Growth and differentiation of a long bone in limb development, repair and regeneration. Dev. Growth Differ. 56, 410–424 (2014).
86.Ury, L. A. et al. Campbell Biology (Pearson, 2021).
87.Davidson, L. A., Keller, R. & DeSimone, D. W. Assembly and remodeling of the fibrillar fibronectin extracellular matrix during gastrulation and neurulation in Xenopus laevis. Dev. Dyn. 231, 888–895 (2004).
88.Moore, S., Keller, R. & Koehl, M. The dorsal involuting marginal zone stiffens anisotropically during its convergent extension in the gastrula of Xenopus laevis. Development 121, 3131–3140 (1995).
89.Engler, A. J., Sen, S., Sweeney, H. L. & Discher, D. E. Matrix elasticity directs stem cell lineage specification. Cell 126, 677–689 (2006).
90.Chen, D. Y., Crest, J., Streichan, S. J.& Bilder, D. Extracellular matrix stiffness cues junctional remodeling for 3D tissue elongation. Nat. Commun. 10, 1–15 (2019).
91.Pickup, M. W., Mouw, J. K. & Weaver, V. M. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 15, 1243–1253 (2014).
92.Elosegui-Artola, A. The extracellular matrix viscoelasticity as a regulator of cell and tissue dynamics. Curr. Opin. Cell Biol. 72, 10–18 (2021).
93.Davies, P. C., Demetrius, L. & Tuszynski, J. A. Cancer as a dynamical phase transition. http://tbiomed.biomedcentral.com/articles/10.1186/1742-4682-8- 30 (2011).
94.Sunyer, R. et al. Collective cell durotaxis emerges from long-range intercellular force transmission. Science 353, 1157–1161 (2016).
95.Duda, M. et al. Polarization of myosin II refines tissue material properties to buffer mechanical stress. Dev. Cell 48, 245–260.e7 (2019).
96.Chanet, S. et al. Actomyosin meshwork mechanosensing enables tissue shape to orient cell force. Nat. Commun. 8, 1–13 (2017).
97.Collinet, C., Rauzi, M., Lenne, P. F. & Lecuit, T. Local and tissue-scale forces drive oriented junction growth during tissue extension. Nat. Cell Biol. 17, 1247–1258 (2015).
98.Hakim, V. & Silberzan, P. Collective cell migration: a physics perspective. Rep. Prog. Phys. 80, 076601 (2017).
99.Fodor, É. et al. Activity-driven fluctuations in living cells. EPL (Europhys. Lett.) 110, 48005 (2015).
100.Ahmed, W. W. et al. Active mechanics reveal molecular-scale force kinetics in living oocytes. Biophysical J. 114, 1667–1679 (2018).
101.Bernheim Groswasser, A., Gov, N. S., Safran, S. A. & Tzlil, S. Living matter: mesoscopic active materials. Adv. Mater. 30, 1707028 (2018).
102.Rupprecht, J. F., Singh Vishen, A., Shivashankar, G. V., Rao, M. & Prost, J. Maximal fluctuations of confined actomyosin gels: dynamics of the cell nucleus. Phys. Rev. Lett. 120, 098001 (2018).
103.Berthier, L., Flenner, E. & Szamel, G. Glassy dynamics in dense systems of active particles. J. Chem. Phys. 150, 200901 (2019).
104.Vicsek, T. & Zafeiris, A. Collective motion. Phys. Rep. 517, 71–140 (2012).
105.Weeks, E., Crocker, J., Levitt, A., Schofield, A. & Weitz, D. Three-dimensional direct imaging of structural relaxation near the colloidal glass transition. Science 287, 627–631 (2000).
106.Pouliquen, O. Velocity correlations in dense granular flows. Phys. Rev. Lett.
93, 1243–1253 (2004).
107.Maxwell, J. C. Xlv. on reciprocal figures and diagrams of forces. Lond. Edinb. Dublin Philos. Mag. J. Sci. 27, 250–261 (1864).
108.Laman, G. On graphs and rigidity of plane skeletal structures. J. Eng. Math. 4, 331–340 (1970).
109.Thorpe, M. Rigidity percolation. in Physics of Disordered Materials. (eds Adler, D., Fritzsche, H. & Ovshinsky, S. R.) 55–61 (Springer, 1985).
110.Thorpe, M. F. Continuous deformations in random networks. J. Non- Crystalline Solids 57, 355–370 (1983).
111.van Hecke, M. Jamming of soft particles: geometry, mechanics, scaling and isostaticity. J. Phys.: Condens. Matter 22, 033101 (2009).
112.Rauzi, M., Verant, P., Lecuit, T. & Lenne, P.-F. Nature and anisotropy of cortical forces orienting Drosophila tissue morphogenesis. Nat. Cell Biol. 10, 1401–10 (2008)
加入自生成结构读书会
生命是有心(意识)的,但当你一层层剥开生命的组织,却只有心脏而没有一颗“心灵”;打开大脑皮层看到一个个神经元,却看不到“意识”。然而,生命与意识都具有“自我生成“的能力,生命自发从非生命中生成,意识自发从生命中生成。更惊人的是,生命与意识的自我生成结构似乎很相似。如果这个假设成立,那么最可能的备选结构会是什么呢?
为了更深入地认识复杂活系统的自生成结构,集智科学家小木球(仇玮祎)联合周理乾、王东、董达、刘宇、苑明理、傅渥成、章彦博等科学哲学、计算机科学、物理学和生物化学等学科的一线研究者共同发起组织《自生成结构系列读书会》。其中已经完结,读书会所形成的自生成结构社群集结了相关领域的教授、硕博及产业界人士。
原标题:《前沿综述:相变怎样塑造生命系统》
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2024 上海东方报业有限公司

