- +1
【重磅综述】线粒体-细胞核通讯在表观和衰老中的作用
线粒体的蛋白质组由细胞核和线粒体DNA (mtDNA)共同编码,维持线粒体与细胞核之间的交流对于线粒体的功能十分重要。衰老过程通常伴随着线粒体的变化,这些变化会影响线粒体与细胞核之间的交流,导致基因表达的变化并影响衰老进程。中国科学院遗传与发育生物学研究所田烨课题组于2022年4月在Trends in Biochemical Sciences上发表了题为“Mitochondrial-to-nuclear Communication in Aging: An Epigenetic Perspective”的综述, 总结了线粒体-细胞核之间的交流信号,深入讨论了多种线粒体代谢物以及线粒体损伤信号通过调节表观修饰参与寿命调控的分子机理,并探讨干预线粒体代谢可以作为一种潜在的延缓衰老的治疗策略。
摘 要
年龄相关的线粒体变化参与衰老过程。除了已知的产能和生物合成功能之外,线粒体可以将它们的状态通过信号交流传递给细胞核,激活转录程序来适应内稳态胁迫,并对于机体健康和衰老有着重要意义。越来越多的研究揭示线粒体到细胞核的通讯通过线粒体代谢物水平的变化或者胁迫信号造成各种表观变化,帮助维持机体内稳态并影响衰老。在这里,我们总结了最近关于线粒体-细胞核通讯调控表观基因组以及调控衰老过程的机制。理解线粒体代谢物如何作为促长寿信号以及衰老如何影响这一通讯过程,将帮助我们发展促进健康和长寿的干预措施。
亮 点
·线粒体-细胞核交流在细胞稳态和衰老过程中发挥重要作用。
·线粒体代谢物是表观修饰的底物或者介导因子。
·线粒体-细胞核胁迫信号通过调控表观修饰调控调节寿命。
线粒体-细胞核通讯在表观基因组和衰老中的作用
线粒体是细胞能量代谢的中心,可以通过三羧酸循环和氧化磷酸化产生ATP。线粒体蛋白质组大部分是由核基因组编码的,只有电子传递链中的13个亚基是由线粒体DNA编码的。因此,线粒体与细胞核之间的交流对于协调线粒体氧化磷酸化复合物的表达、翻译和组装至关重要。在衰老过程中,线粒体功能下降与线粒体氧化磷酸化活性下降、三羧酸循环相关代谢酶的水平改变、线粒体DNA突变累积、活性氧产生增加、线粒体蛋白质内稳态失调密切相关。线粒体功能失调可能会破坏线粒体与细胞核之间的交流,导致与衰老相关的基因表达变化。
线粒体功能是由核编码基因通过正向信号调控的,通过促进线粒体生物合成或增加线粒体活性来适应细胞的需求。这种调控主要依赖于细胞核编码的转录因子,例如PGC1α、NRF1和其他调控因子。与之对应的是由线粒体到细胞核的反向信号传递。反向信号传递响应各种线粒体异常(蛋白内稳态胁迫、能量缺乏、活性氧增加等)并激活转录重编程以适应代谢(图1)。在参与线粒体与细胞核之间信号交流过程的主要调控因子中,只有SIRT1、AMPK和ATFS-1被研究的比较深入。越来越多的研究表明线粒体代谢物和其他的胁迫信号可以作为信号分子调节表观修饰,调控代谢相关基因的表达从而调节细胞内稳态以及衰老。并且衰老本身也伴随着各种复杂的表观和代谢变化。因此,线粒体和细胞核之间的交流组成了复杂的调控网络使得细胞可以响应时刻变化的代谢环境和衰老相关的胁迫。这篇综述主要讨论了线粒体代谢物和胁迫信号是如何通过影响表观修饰调控衰老或者促进长寿,并总结了现在对于线粒体代谢物可能作为促长寿因子的研究状况。
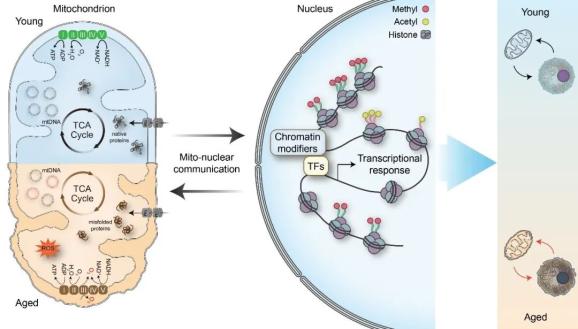 图1 线粒体-细胞核交流与衰老
图1 线粒体-细胞核交流与衰老线粒体代谢物和表观基因组相互作用调控寿命
线粒体除了可以给细胞提供能量之外,也作为一个代谢中心,负责许多大分子物质(如核苷酸、脂类和蛋白质)的合成。一般认为来源于线粒体的代谢中间产物是细胞代谢的副产物,但是现在人们逐渐认识到来源于线粒体的代谢信号可以通过非代谢机制调节细胞核中的表观修饰(图2)。线粒体代谢物和表观基因组之间的互作可以改变细胞核基因表达,从而调控细胞内稳态和衰老过程。在这一部分,我们将主要讨论线粒体代谢物(如乙酰辅酶A、α-酮戊二酸、NAD+和甲硫氨酸)作为可能的长寿调控因子以及这些代谢物丰度的改变如何影响表观基因组和衰老过程。
乙酰辅酶A
三羧酸循环(也称柠檬酸循环或克氏循环)通过氧化来源于碳水化合物、脂肪和蛋白质的乙酰辅酶A释放储存的能量。在线粒体中,乙酰辅酶A进入三羧酸循环并产生柠檬酸和草酰乙酸。来源于三羧酸循环的中间产物可以被转运到细胞质合成其他物质。例如,柠檬酸可以通过线粒体柠檬酸转运蛋白SLC25A1离开线粒体并在细胞质和细胞核中被ATP-柠檬酸裂合酶(ACLY)转变成乙酰辅酶A和草酰乙酸。此外,在营养限制条件下细胞质中的酰基辅酶A合成酶ACSS2可以利用乙酸盐产生乙酰辅酶A。细胞质的乙酰辅酶A可以被用于合成脂肪酸、类固醇或某些氨基酸。除了在代谢和生物合成中的关键作用之外,乙酰辅酶A也具有传递信号的功能,因为它可以为蛋白乙酰化(如组蛋白乙酰化)提供乙酰基团。乙酰化中和了赖氨酸残基上的正电荷,使得染色质结构开放,促进转录因子结合并影响基因表达。在衰老过程中,各种线粒体胁迫或者病理情况都会改变乙酰辅酶A的丰度以及在亚细胞组分中的分布情况。因此,乙酰辅酶A作为一个二级信使从线粒体传递信号到细胞核来调控代谢适应过程,并可能作用于衰老过程。
虽然乙酰辅酶A是合成代谢和分解代谢的核心代谢物,但是由于其不稳定的特性,对于衰老过程中乙酰辅酶A水平变化的研究仍然很少。在衰老以及衰老相关的神经退行性疾病中,许多线粒体相关功能都会受损。一项研究报导在衰老的小鼠大脑中乙酰辅酶A水平下降,而使用两种小分子化合物CMS121或J147可以恢复大脑中的乙酰辅酶A水平,并伴随着组蛋白H3特定位点乙酰化水平和记忆增强。虽然衰老过程中整体线粒体功能下降,但是在果蝇中的研究惊奇地发现乙酰辅酶A和柠檬酸水平在中年时期有所上升,敲降ACLY降低乙酰辅酶A水平并导致代谢变化和寿命延长。因此,乙酰辅酶A对于寿命调控的影响在不同物种或者不同组织之间并不完全一致,最近的研究表明乙酰辅酶A的区域化以及其在特定组织中对于基因表达的影响是其调控寿命的关键。
在酵母中,抑制乙酰辅酶A进入线粒体会导致乙酸盐在细胞质中累积,过度激活乙酰辅酶A合成酶Acs2p并促进组蛋白乙酰化,导致寿命缩短。Pouikli和其同事观察到在间充质干细胞中乙酰赖氨酸信号随着衰老有一个出乎意料的定位改变,从细胞核转移到线粒体中。他们发现在衰老的间充质干细胞中由于柠檬酸载体水平下降,乙酰辅酶A被困在线粒体中,导致组蛋白乙酰化下降以及衰老相关的染色质凝集。通过外源表达柠檬酸载体或者补充乙酸盐来恢复细胞质乙酰辅酶A水平可以重塑染色质结构并且使间充质干细胞年轻化。这项研究表明线粒体到细胞核的信号交流在干细胞衰老过程中发挥重要作用。
由于在不同物种、不同组织中组蛋白乙酰化随着衰老而变化,未来关注于乙酰辅酶A的分布和相对丰度的研究可能会对线粒体代谢物和组蛋白修饰之间的联系导致的基因表达变化以及衰老相关表型产生更多的见解。
α-酮戊二酸
α-酮戊二酸是三羧酸循环中异柠檬酸脱氢酶IDH1/2通过氧化脱羧作用催化异柠檬酸产生的中间代谢物。α-酮戊二酸也是2-氧戊二酸依赖性双加氧酶(2-OGDDs)的共底物。2-OGDDs是一大类保守的负责催化不同底物(如蛋白、脂类、核酸和中间代谢物)的羟基化反应的酶。此外,因为α-酮戊二酸是一些染色质修饰酶(如组蛋白去甲基化酶JMJDs和DNA去甲基化酶TETs)的底物,因此它在生理过程中通过表观调控发挥着多种作用。值得注意的是,JMJDs和TETs的活性依赖于细胞中α-酮戊二酸与琥珀酸或其他抑制因子(如延胡索酸或2-羟基戊二酸)的比率。因此,线粒体代谢物α-酮戊二酸的水平变化可以通过影响DNA和组蛋白甲基化改变基因表达。
在衰老过程中,线粒体功能和代谢水平逐渐下降,使得α-酮戊二酸愈加不足。有研究报导α-酮戊二酸不足在干细胞中随着年龄而增加。例如,在衰老人类的卵泡液中,α-酮戊二酸水平下降;而在小鼠中补充α-酮戊二酸可以维持卵巢功能。在线虫、果蝇和小鼠中的研究表明补充α-酮戊二酸可以延长寿命。在线虫中,α-酮戊二酸水平在长寿的线粒体突变体中显著增加。补充α-酮戊二酸或者其衍生物I-2-hydroxyglutaratI(R)-2HG可以通过抑制ATP合酶活性延长线虫寿命。在果蝇中,α-酮戊二酸通过激活AMPK信号和抑制mTOR通路延长寿命。此外,在年老的小鼠中以一种钙盐的方式(CaAKG)补充α-酮戊二酸可以促进更长和更健康(炎症因子水平降低)的寿命。
这些研究表明α-酮戊二酸可能是促进人类长寿的一个理想的候选因子,然而α-酮戊二酸对于衰老过程中表观修饰的作用仍然不清楚。最近的一项由Ponce de Leon Health公司资助的研究表明,42个人在服用了Rejuvant (一种基于α-酮戊二酸的药物)4-10个月可以导致其生物学年龄有近8岁的逆转。他们的结论主要基于分析DNA甲基化时钟(一种确认的衰老生物标记)。然而,补充α-酮戊二酸会导致基因组上一些CpG位点的去甲基化和过度甲基化,说明其对于衰老过程中的甲基化有着更广泛的影响,例如其他的代谢功能。值得注意的是,过表达H3K27去甲基化酶JMJD-1.2/PHF8和JMJD-3.1/JMJD3 (α-酮戊二酸是其可能的底物)可以诱导线粒体基因的表达并延长线虫寿命。未来通过检测染色质甲基化图谱来研究补充α-酮戊二酸是否可以减慢衰老的表观时钟将会是一个有趣的研究方向。
NAD+
NAD+及其还原形式NADH是与线粒体产能紧密相关的重要代谢物。NAD+是氧化还原反应的辅酶,对于核心代谢通路(如三羧酸循环、电子传递链、糖酵解和脂肪酸β氧化)至关重要。细胞中NAD+/NADH的比率调节酶的活性(如组蛋白去乙酰化酶Sirtuins、cADP-核糖合成酶CD38和多聚ADP-核糖聚合酶PARPs)并影响基因表达。在酵母、线虫和小鼠上的研究表明,NAD+和sirtuins与衰老调控相关。在衰老过程中,小鼠不同组织的NAD+水平下降。人类大脑中NAD+水平也随着年龄下降。对于寿命延长与补充不同NAD+前体之间的联系已经在不同物种中进行了大量的研究。对于NAD+在代谢和衰老中的作用在其他综述中已经被描述过了。
Sirtuins感受胞内NAD+浓度并通过蛋白去乙酰化传导信号,其在寿命调控中的作用最初是在酵母中被报道的。然而值得注意的是,在线虫和果蝇中,Sir2在衰老中的功能都存在争议。Sir2过表达诱导的长寿后来发现是由于转基因的脱靶效应和二次突变导致的。哺乳动物中有七个Sirtuins,SIRT1是与酵母中Sir2最类似的。在小鼠中过表达Sirt1可以阻止衰老相关的代谢下降但是不能延长寿命。PARP (一种依赖于NAD+的酶)参与DNA修复、炎症反应和细胞死亡,其在代谢和衰老中的作用,以及与sirtuins的竞争关系已经在最近的其他综述中讨论过了。
作为一个核心代谢调控因子,NAD+帮助维持线粒体功能以及细胞内稳态。在早衰疾病模型中,NAD+可以通过介导线粒体自噬和线粒体合成促进线粒体功能。通过增强线粒体复合物I活性来再生线粒体NAD+可以延长大脑中线粒体复合物I缺陷小鼠的寿命。在衰老过程中维持其水平也可以保持线粒体与细胞核之间的通讯。在线虫中,用遗传手段或者药物恢复NAD+水平可以通过SIR-2.1激活线粒体未折叠蛋白反应(UPRmt)来延长寿命。最近的一项研究也表明在过氧化氢诱导的氧化胁迫下,线粒体NAD+可以通过控制细胞核ADP-核糖基化来促进线粒体与细胞核之间的通讯。因此,通过调控NAD+水平来促进线粒体-细胞核交流可能可以作为一种延缓衰老相关代谢衰退的措施。
甲硫氨酸
传统观点认为氨基酸只是组成蛋白质的基石,但是氨基酸也可以被分解成三羧酸循环的中间产物进入线粒体产能,并且在信号转导过程中也发挥着重要作用。从酵母到人类中,氨基酸的浓度都随着衰老而改变,并且在长寿物种中也有所改变。在这一部分,我们用甲硫氨酸作为一个例子来讨论氨基酸影响衰老过程背后的机制。
除了在翻译起始过程中的作用之外,甲硫氨酸也参与多种代谢途径,包括甲硫氨酸循环、反式硫化途径和多胺合成。甲硫氨酸是S-腺苷甲硫氨酸(SAM)的主要氨基酸来源,SAM是甲基化的甲基供体。在细胞质中,叶酸循环与甲硫氨酸循环偶联来产生SAM,线粒体一碳循环也可以维持SAM。细胞质的SAM水平决定了组蛋白甲基化水平,特别是H3K4me3来维持特定的细胞状态。因此,线粒体代谢波动诱导的SAM水平变化可能可以影响组蛋白和DNA甲基化,从而影响细胞内稳态和寿命。
甲硫氨酸限制可以延长许多物种的寿命。在小鼠中,饮食限制甲硫氨酸可以改善代谢健康并且延缓衰老相关疾病。在酵母中,甲硫氨酸限制通过一种反向信号反应来改变线粒体功能,诱导长寿。在线虫中,敲降催化SAM合成的甲硫氨酸腺苷基转移酶基因sams-1可以促进寿命延长。一些研究表明甲硫氨酸在寿命调控中起着负调控作用。然而,一些长寿的果蝇株系含有更高水平的甲硫氨酸。此外,饮食限制诱导的生育能力下降可以通过补充甲硫氨酸来恢复,而不抑制饮食限制诱导的长寿。在果蝇中的研究表明,在衰老过程中,甲硫氨酸代谢发生重编程,伴随着S-腺苷同型半胱氨酸(SAH)水平上升,SAH是由SAM转变而来的。抑制SAH水解酶dAhcyL1活性可以降低SAH水平,抑制H3K4三甲基化,从而延长健康寿命。这些结果表明甲硫氨酸在寿命调控中起着有益的作用,说明甲硫氨酸代谢有可能比甲硫氨酸的绝对水平更加重要。
亚精胺(spermidine)是一种来源于甲硫氨酸的自然多胺,在从酵母到哺乳动物中,外源补充亚精胺可以延长寿命,而没有明显的副作用。亚精胺通过抑制组蛋白乙酰转移酶HAT激活了组蛋白H3去乙酰化,从而上调了自噬相关转录并诱导自噬。由于内源亚精胺水平与年龄呈负相关,并且在一项最近的临床实验中,补充亚精胺表现出了很好的安全性,因此富含亚精胺的饮食有极大潜力可以促进长寿和健康衰老。综上所述,这些研究表明饮食氨基酸干预可能可以通过独立于它们传统的蛋白代谢功能的表观调控重塑线粒体到细胞核的交流来影响细胞内稳态和衰老。
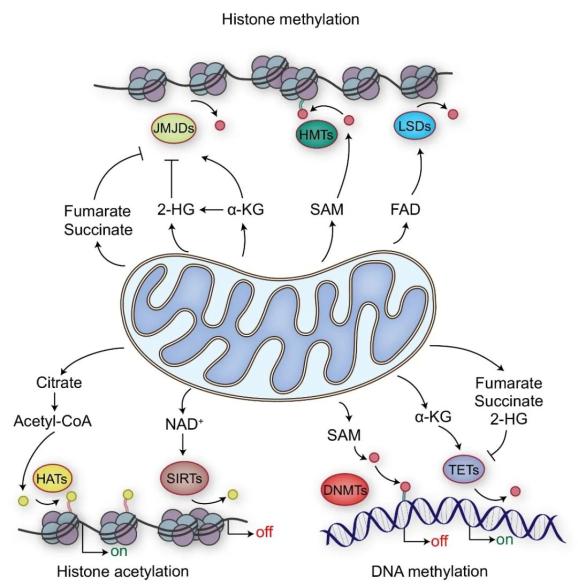 图2 线粒体代谢物与表观修饰
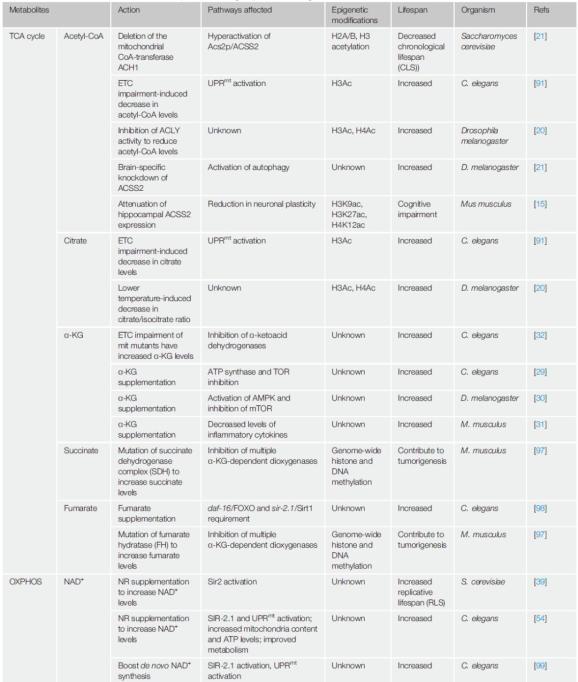
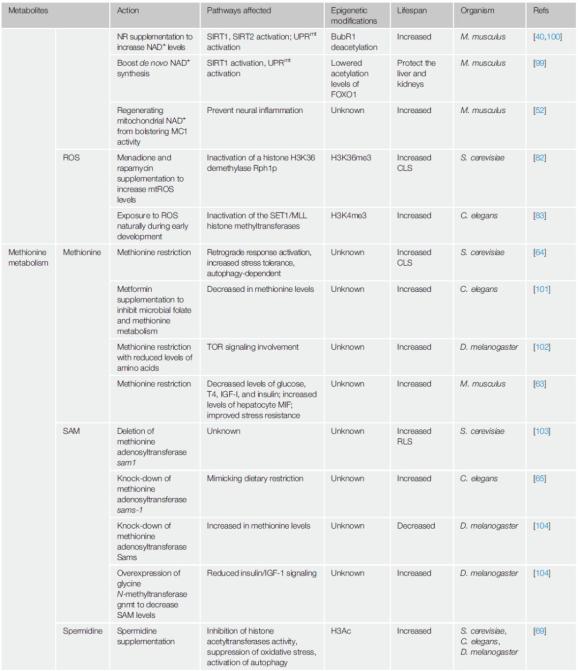
图2 线粒体代谢物与表观修饰表1 不同物种中与寿命相关的线粒体代谢物
 线粒体到细胞核的胁迫信号通过表观调控决定寿命
线粒体到细胞核的胁迫信号通过表观调控决定寿命细胞的代谢状态不止在衰老中改变,也易受到环境刺激的影响。线粒体受到干扰可以诱发胁迫信号传导,将信号传递给细胞核从而帮助适应。线粒体功能受损时(如线粒体DNA丢失,线粒体DNA突变累积,呼吸作用受损,线粒体蛋白稳态失调,活性氧产生等),不同的通路会被激活。这种信号促进线粒体与细胞核之间的交流来改变基因表达,导致代谢适应和长寿。越来越多的研究已经揭示了线粒体胁迫导致的染色体修饰变化促进了线粒体与细胞核交流,从而留下了一种表观记忆,可能可以影响衰老过程。在这一部分,我们主要讨论线粒体胁迫信号如何通过表观调控调节衰老。
线粒体活性氧(mtROS)
活性氧是正常有氧代谢的副产物。许多年来,活性氧都被认为只是导致生物大分子(如DNA,蛋白质和脂类)氧化损伤并诱导细胞氧化胁迫响应和加速衰老的有害产物。氧化还原信号通过调节转录因子、代谢酶的活性和表观修饰调控细胞内稳态。然而,氧化胁迫响应在多大程度上参与衰老过程,是作为原因、结果,还是仅仅只是相关性,仍然不清楚。
线粒体是活性氧产生的主要来源,特别是自由基,例如超氧化物。衰老与线粒体酶活性下降以及活性氧产生增加相关。一项研究表明,在成虫第三天的线虫中,线粒体中随机激增的超氧化物产生与寿命负相关(除长寿的线粒体突变体外)。然而,在长寿的裸鼹鼠中,活性氧的产生水平与小鼠类似。mtROS曾经被认为仅仅只是细胞内稳态的破坏者,造成氧化损伤和加速衰老。然而后续的许多研究表明mtROS可以作为重要的信号分子调控衰老、炎症和癌症。
从酵母到啮齿动物的研究表明,mtROS信号可以通过呼吸抑制、卡路里限制、抑制TORC1信号传递、适度低氧暴露或温度胁迫来促进胁迫抵抗和寿命延长。但是很少有研究鉴定到其中传递信息来激活高度特异的转录程序的信号。mtROS信号可以通过两个DNA损伤响应的激酶激活一个JMJD组蛋白H3K36去甲基化酶Rph1p,增强沉默蛋白Sir3p的结合,抑制亚端粒区域的转录来延长酵母的时间寿命。在线虫中,发育过程中增加的ROS水平会增强生命后期的胁迫抗性并促进寿命延长。早期的ROS介导的抑制SET1/MLL组蛋白甲基转移酶导致整体H3K4me3水平下降,增强氧化还原内稳态并最终促进长寿。因此,线粒体功能失调或者mtROS信号传递可能可以影响寿命,并且发生的时间以及ROS产生的水平都是重要因素。
UPRmt
线粒体功能由一系列感知线粒体功能失调并响应细胞代谢需求的质量控制途径监控。这些胁迫响应是由线粒体中产生的信号启动的,并诱导细胞核中的响应来保护线粒体功能。虽然严重的线粒体胁迫是有害的,但是发育过程中适当的线粒体胁迫可以通过表观调控对机体的寿命产生有益效应。
在线虫中,发育过程中用RNAi敲降电子传递链组分不仅可以延长寿命,而且可以激活一种称为UPRmt的转录响应,从而缓解线粒体中的蛋白折叠压力。有趣的是,激活UPRmt对于线粒体胁迫诱导的长寿是必需的,并且激活UPRmt的能力在衰老过程中显著下降。线虫中UPRmt的主要调控因子是转录因子ATFS-1,它具有一个线粒体定位信号和一个核定位信号。当线粒体功能受损时,ATFS-1的线粒体转运效率下降,从而使得ATFS-1在核内累积,激活UPRmt。ATFS-1自身代表了一种胁迫状态下的线粒体到细胞核的交流途径。值得注意的是,严重的线粒体胁迫(例如抑制线粒体转运机器)可以很强地诱导UPRmt但是不能延长线虫寿命。此外,ATFS-1的功能获得性突变体也并不长寿,说明UPRmt的组成性激活并不足以诱导寿命延长。
值得注意的是,发育过程中只有一段时间窗口可以产生有益效应,说明早期发育过程中的线粒体功能失调可以改变表观基因组,从而改变机体的生理过程,最终影响寿命。的确,线粒体胁迫会造成广泛的染色质重塑,通过多种表观因子诱导UPRmt。在线粒体胁迫发生时,ATF7IP/LIN-65在核内累积,该过程需要组蛋白H3K9甲基转移酶MET-2/SETDB1核同源框转录因子DVE-1/SATB1来促进染色质紧缩。与此同时,JMJD组蛋白去甲基化酶JMJD-1.2/PHF8和JMJD-3.1/JMJD3也对于线粒体胁迫诱导的UPRmt激活和寿命延长是必需的。然而,线粒体胁迫诱导的表观改变独立于ATFS-1,说明来源于线粒体的其他胁迫信号对于染色质重塑是必需的。
朱頔等人发现线粒体胁迫导致的乙酰辅酶A水平的下降作为一种信号促进核小体重塑和组蛋白去乙酰化酶(NuRD)复合体介导的染色质重塑和寿命延长。NuRD复合体中的组分,组蛋白去乙酰化酶HDA-1与DVE-1协同调控UPRmt响应。其哺乳动物中的同系物HDAC1/2也在调控线粒体内稳态上起着保守的作用。NuRD组分缺失足以造成衰老相关表型的发生。值得注意的是,线粒体胁迫可以促进NuRD组分在核内累积,过表达NuRD复合物组分就足以诱导线虫寿命延长。此外,一个乙酰转移酶CBP-1/p300正调控UPRmt,并且对于线粒体胁迫诱导的长寿是必需的。
可以推测NuRD复合物介导的组蛋白去乙酰化可能和LIN-65/MET-2介导的组蛋白甲基化协同作用,促进染色质紧缩,而JMJD组蛋白去甲基化酶和CBP-1可能负责维持染色质特定位点的开放构象来促进UPRmt的诱导。因此,染色质修饰可以响应线粒体损伤,促进线粒体到细胞核的通信,从而留下影响衰老过程的表观遗传印记(图3)。未来的研究需要解决线粒体干扰导致的其他线粒体代谢物水平的变化是否可以重塑表观基因组来调控寿命。UPRmt最初是在哺乳动物细胞中被鉴定发现的,在线虫中,其分子机制得到了深入的研究。在哺乳动物中,UPRmt的诱导是否需要表观调控或者发育过程中的线粒体功能失调是否可以促进长寿仍待研究。胁迫响应发生的时间以及程度,或者线粒体干扰后其他的信号通路,都是决定寿命的关键因素。
线粒体胁迫不仅仅可以传递信号给同一个细胞中的细胞核,还可以被其他组织或器官感知到,来协调整个机体以应对线粒体功能失调,这对于机体内稳态和衰老十分重要。在未来,对于系统性协调线粒体到细胞核的胁迫信号网络调控寿命的研究会是一个重要的研究方向。
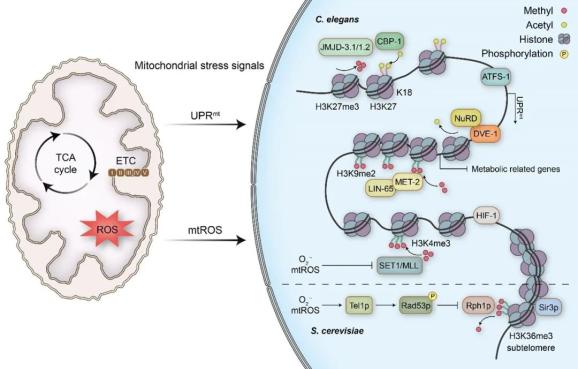 图3 线粒体到细胞核的胁迫信号传导通过表观修饰调控寿命
图3 线粒体到细胞核的胁迫信号传导通过表观修饰调控寿命框1 系统性控制线粒体到细胞核的胁迫信号调控寿命
来源于一个组织的线粒体胁迫信号可以通过一种细胞非自主性的调控方式诱导远端组织的胁迫响应,使得机体可以更好地适应衰老过程中的局部线粒体胁迫。肌肉特异性敲除线粒体细胞色素c氧化酶COX造成一系列疾病表型并缩短小鼠寿命。携带有线粒体转录因子TFAM缺陷的T细胞表现为伴随衰老的慢性诱导炎症,并最终导致多重病症和早衰。到目前为止,不同组织中的线粒体缺陷在衰老过程中如何协调仍然不清楚。
在线虫神经中的电子传递链损伤可以通过一种细胞非自主性的方式激活肠道的UPRmt并延长寿命。有假说认为分泌性的线粒体细胞分子(mitokine)信号从具有线粒体损伤的组织中传递出来激活远端组织中的线粒体胁迫响应。进一步的研究表明,retromer介导的Wnt信号作为mitokine信号参与细胞非自主性诱导UPRmt。神经特异性过表达Wnt配体EGL-20就足以诱导肠道中的UPRmt和寿命延长。有趣的是,神经的线粒体胁迫也可以被生殖腺中的线粒体感知并作出反应,以一种依赖于Wnt信号的方式来促进母系遗传的多代的mtDNA水平上升。UPRmt的跨代遗传和mtDNA的升高使得后代线虫可以活的更长,并且对胁迫有更强的抗性,同时付出的代价则是发育减慢和生育力下降。
一项利用不同野生型线虫的研究表明神经肽介导的胶质细胞-神经元信号传导的自然遗传多样性以一种依赖于SIR-2.1介导的UPRmt的方式调控衰老速率。在果蝇的肌肉细胞中干扰线粒体功能可以激活UPRmt并且诱导ImpL2(胰岛素结合蛋白)信号来抑制胰岛素类似生长因子信号通路(IIS)并延长寿命。在哺乳动物中,系统性协调线粒体胁迫响应也是保守的。在小鼠中,下丘脑POMC神经元中的线粒体功能失调会导致更高的代谢更新速率、肥胖、重塑脂肪代谢。线粒体疾病的病人表现为肌无力,并且血浆中有更高水平的FGF-21和GDF-15。在高等动物中,哪些组织器官负责协调机体的促长寿信号将会是重要的研究方向。
悬而未决的问题
线粒体代谢物的产生如何在时间和空间上受调控来响应线粒体功能失调诱导表观变化?
参与线粒体-细胞核通讯的特定表观因子是什么?它们如何与转录因子协同响应不同的外部或内部刺激?
不同的线粒体代谢物是单独作用还是协同作用于表观基因组来调控衰老进程?某些器官或组织在衰老过程中是否更易于在维持线粒体-细胞核通讯方面处于风险?
线粒体-细胞核通讯的干预措施能否模拟有益的表观效应来延缓衰老或者减轻衰老相关疾病?
结论与展望
本文主要总结了线粒体到细胞核的交流在调控衰老过程中的重要作用。外在或内在的代谢因素可以影响线粒体功能,从而通过表观修饰改变基因表达。通过操控一些核心线粒体代谢物来促进细胞代谢和逆转衰老相关表观变化可能可以称为延缓衰老的临床策略。
越来越多的证据表明线粒体代谢在时间和空间上都受到紧密调控来引发对于营养水平和信号因素的响应。组织特异性或者线粒体特异性代谢组分析将使我们对于代谢物导致的表观调控有更进一步的理解。细胞非自主性线粒体交流的信号转导是如何被调控的仍然不是十分清楚。组织和器官之间的互作来调控机体的代谢状态也可能成为相关疾病的治疗策略。
饮食对于代谢和寿命有着十分重要的影响。一种公认的能促进长寿的饮食干预是饮食限制。有证据表明饮食限制诱导的寿命延长可以被代谢物的水平变化所影响的DNA甲基化介导。然而,饮食限制的有益效应在晚年小鼠中不能被有效诱导。发育过程中的线粒体胁迫造成染色质结构的广泛改变来促进UPRmt,使得早期的胁迫响应可以导致寿命延长。这一证据说明,某种程度上,表观变化是可逆的以及染色质是动态变化的。因此,适当的干预饮食中的营养成分可能是一种健康有效的方式来调控线粒体功能并延缓衰老。
尽管线粒体作为信号转导中心的重要性不言而喻,但是目前我们对线粒体到细胞核通讯信号的理解还十分有限。线粒体代谢物的水平和分布在空间和时间上是如何被调控的?不同组织和器官之间是如何相互交流线粒体代谢水平,共同协调机体整体的代谢和衰老过程的?在今后,组织特异的线粒体代谢组学分析将会引导我们深入理解线粒体代谢物驱动表观遗传的调控机制。线粒体代谢物如何以组织特异的方式影响基因组上特定位点的表观修饰也是该领域未来值得关注的重要问题。开发有关干预线粒体到细胞核通讯的方式,以促进细胞代谢或逆转与年龄相关的表观遗传变化,也将成为未来延缓衰老或衰老相关疾病的方向之一。

原标题:《【重磅综述】线粒体-细胞核通讯在表观和衰老中的作用》
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2026 上海东方报业有限公司

