- +1
生物演化史上的“黑洞”,可能需要物理学来填补
科学之精要在于能够提供规律,比如物理学中最迫切的问题在于,为什么物理定律是这样的?使用什么样的基本规律才能推演出宇宙中已知的各种物理特性?生物学的规律性远弱于物理学,尽管生物学规律不需要像物理定律那样严密普适,演化生物学的预言能力还是弱得令人汗颜。关于演化的分子生物机制和地球的生命历史,我们已经积累了海量的知识,但对其中的规律性仍然知之甚少。在生命的历史中,哪些部分是出于偶然,哪些部分又有物理定律和约束条件决定?
撰文 | Nick Lane
译者 | 严曦
在生物学的核心地带,存在着一个未知的黑洞。坦白说,我们不知道生命为什么是现在这样。地球上所有的复杂生命拥有一个共同祖先,它从简单的细菌演化而来,在40亿年的漫长岁月中只出现了一次。这究竟是一个反常的孤立事件,还是因为其他的复杂生命演化“实验”都失败了?我们不知道。已知的是,这个共同祖先一出场,就已经是一个非常复杂的细胞。它的复杂程度,与你身上的细胞不相上下。这份复杂性遗产传给了你我,也传给了其他所有后代,从树木到蜜蜂。你可以试试在显微镜下观察自己的细胞,并和蘑菇细胞比较。二者几乎无从分辨。我们与蘑菇的生命显然天差地远,那么,为什么我们的细胞却如此相似?而且并不只是外观相似,所有复杂生命都有同一套细胞特征,从有性生殖到细胞衰老再到细胞凋亡,其机制之精巧复杂,与物种间的相似程度同样惊人。为什么这些特征会在我们的共同祖先身上积聚?为什么在细菌身上却找不到这些特征独立演化的痕迹?如果这些特征是通过自然选择微步演化而来,每一小步都带来一点点优势,那么为什么类似的特征没有出现在各类细菌中?对这些问题的探讨可谓众说纷纭,但学界至今没有给出令人信服的解释。
这些问题反映了地球生命奇特的演化轨迹。生命在地球形成约5亿年后就已出现,距今大约40亿年。然而,此后的20多亿年中,也就是地球历史一半的时间,生命一直停滞在细菌水平。直到40亿年后的今天,细菌仍然保持简单的形态,虽然它们发展出丰富的生物化学代谢能力。所有形态复杂的生物,包括植物、动物、真菌、藻类和阿米巴原虫等单细胞原生生物,与细菌形成了鲜明的对比。它们是同一个祖先的后代,这个祖先大约于20亿~15亿年前出现,从外形到内在都是一种“现代”细胞,拥有精细的内部结构和空前的能量代谢水平。所有这些新特征,都由一套复杂的蛋白纳米机器驱动,由数以千计的新基因编码,而这些基因在细菌身上几乎从未发现。在复杂生命的共同祖先与细菌之间,没有现存的演化中间型,没有“缺失环节”来揭示为什么这些复杂的特征会出现,以及它们是如何演化的。在细菌的简单与其他一切生命令人敬畏的复杂之间,只有一片无法解释的空白。一个演化的黑洞。
人类为什么会遭到各种疾病的侵袭,这是一系列复杂到无法想象的问题。为了寻求答案,人类每年都会在生物医学研究上投入巨额的金钱。我们现在对基因和蛋白质的关系、对生物调节系统之间的反馈互动,都掌握了海量的细节。我们建立了精密的数学模型,设计了计算机模拟程序,以信息化的方式来重建这些生物过程。然而我们仍然不知道,所有这些生物组件都是如何演化而来的!如果我们不知道细胞为什么是这样运行的,又如何能指望理解疾病呢?不了解历史,我们就不可能理解一个社会;不了解细胞的演化史,我们就不可能理解细胞的运作方式。这些问题不仅有重要的实用意义,它们本身也是人类要面对的终极问题:我们为什么会存在?是什么样的法则创生了宇宙、恒星、太阳、地球,以及生命本身?同样的法则是否也在宇宙中的其他地方创造了生命?外星生命是否和我们相似?诸如此类的形而上之问,关乎我们何以为人的核心。然而,自人类发现细胞350年之后,我们仍然不知道地球生命为什么会是这样。
读者你可能还没注意到人类在这方面的无知,这不是你的错。各种教科书和学术刊物上满载科学信息,但绝大多数都不会探究这种“幼稚”的问题。我们被互联网上各式各样、良莠不齐的信息淹没,难以辨别它们的真伪与用途。但这不仅仅是信息过载的后果,就连生物学家自己,对这个专业领域中心的黑洞,也没有多少清醒的认识。绝大多数生物学家忙于研究其他问题。大多数学者研究大型生物、特定的动物或植物,少数人研究微生物,更少数的人研究细胞的早期演化。另外,生物学界还要担心神创论者和智慧设计论的攻击。他们担心:承认科学家不知道所有的答案,也许会让反演化论者乘虚而入,让他们嘲笑自己对演化实际上一无所知。这种担心其实毫无必要,我们对已经掌握的知识很有信心。生命起源和早期细胞演化理论能够解释海量的事实,与其他科学知识严密符合,能对未知的生物学关系进行预测,并且经得起实证检验。我们对自然选择机制,对另一些塑造基因组的随机过程都理解得非常充分。所有这些事实,都能与细胞的演化理论互相印证。然而,我们对事实的高度掌握恰恰凸显了一个问题:我们不知道为什么生命会以如此奇特的路径演化。
科学家是充满好奇心的人,如果这个问题确实像我说的这么严峻,它本应该广为人知。然而实际情况并非如此,科学界对这个问题的意识远没有明确。现有的各种解释互相争论,但大都晦涩难懂,反而掩盖了问题本身。另一个难点在于,问题的相关线索分散在众多不同的研究领域之中:生物化学、地质学、种系发生学、生态学、化学和宇宙学。几乎没有人能成为所有这些领域真正的专家。生物学现在正处于“基因组革命”的进程之中,我们掌握了成千上万种生物完整的基因组序列,包含亿万数位的代码。在这些数字化的生物信息中,从远古遗留至今的线索经常互相矛盾,令人迷惑。对这些数据的解读要求研究者拥有精深的逻辑、计算和统计技能,生物学上的理解反倒被挤到了从属地位,“有当然好,没有也无大碍”。大数据研究方法的迷雾在我们周围涌动,争论交缠。每当稍稍拨开迷雾,更加离奇的新课题就会涌现。科学家曾经的自信从容逐渐消失,我们现在面对着一幅崭新的生物学场景:真实、严峻、令人不安。然而,从一个研究者的角度来看,能够发现一个重要的新课题并寻求答案,非常激动人心!生物学中最大的问题还有待解决,本书正是我起手的尝试。
细菌和复杂生命有什么样的关系?早在17世纪70年代,荷兰科学家安东尼·列文虎克(Antonie van Leeuwenhoek)在自制的显微镜下发现微生物时,对这个问题的初步研究就开始了。他镜头下生机勃勃的“微型动物”,让当时很多人都难以置信。但这些生命的存在,很快被同样天才的罗伯特·胡克(Robert Hooke)再次证实。列文虎克还发现了细菌,他在1677年那篇著名的论文中写道:它们“小到令人难以置信;以我视野中能看到的估计,100个这样极其微小的动物排成一行,宽度也超不过一粒细砂;如果我的估计正确,那么一百万个加起来,也不会比一粒细砂重”。不少后世的研究者怀疑,列文虎克是否真的用他简单的单镜头显微镜观察到了细菌。现在已经没有争议:他确实做到了。可以通过两点证明:首先,他发现细菌无处不在——雨水中,海水中,并不仅仅在自己的牙齿上才能找到;其次,他通过敏锐的直觉发现了两种微生物的区别,一种被他称为“非常微小的动物”(即细菌),另一种则是“巨大的怪物”(实际是单细胞的原生生物,protists)。他区别二者的依据,在于观察到后者旺盛的运动行为和“小脚”(纤毛);他甚至注意到有些较大的细胞由多个小“液泡”组成,并与细菌进行了比较(当然他用的不是这些现代术语)。几乎可以肯定,列文虎克在这些液滴中观察到了细胞核,即所有复杂细胞贮藏基因的地方。接下来,关于这个课题的研究停滞了几个世纪。在列文虎克发现微生物50年后,著名的分类学家卡尔·林奈(Carl Linnaeus)把所有微生物不加区别地分在Vermes门(意为“蠕虫”)的Chaos属(意为“无形状”)。到了19世纪,与达尔文同时代的德国演化论学者恩斯特·海克尔(Ernst Haeckel)才再次确认微生物之间存在重大差别,并把细菌与其他微生物区分开来。20世纪中期以前,关于细菌的生物学认知没有多少进展。
生物化学的统一,使细菌研究迈入了一个关键阶段。此前,细菌花样百出的新陈代谢能力让它们似乎无法归类。它们可以依靠任何物质生存,从混凝土到电池液到气体。如果这些完全不同的生活方式没有任何共同点,我们用什么依据来确定细菌的分类呢?如果无法归类,我们又怎能理解细菌呢?正如化学元素周期表为化学研究带来了理论的统一,生物化学也为细胞演化研究带来了秩序。另一位荷兰人阿尔贝特·克吕沃尔(Albert Kluyver)阐明:生命令人眼花缭乱的多样性,其实是由非常相似的生物化学过程支撑。呼吸作用、发酵作用和光合作用虽然各有区别,但全都有共同的生物化学基础。这种概念的整合,证明了所有生物起源于一个共同的祖先。克吕沃尔认为,在生物化学层面,对细菌成立的理论,对大象也成立;细菌和复杂生物之间几乎没有界限。细菌的代谢多样性远超复杂生物,但二者最基本的生命活动过程都很相似。克吕沃尔的学生科内利斯·范尼尔(Cornelis van Niel),以及罗杰·斯塔尼尔(Roger Stanier),可能是那个时代最能理解细菌的本质不同于其他生物的科学家。他们认为,细菌就像原子,不可再分。它们是最小的生物功能单位。很多细菌和人类一样能进行有氧呼吸,但必须通过整个细菌才能执行这种功能,细菌没有人类细胞中专门进行呼吸作用的组件。细菌繁殖时会分裂,但从功能角度来说,细菌不可再分。
过去的半个世纪中出现了三次颠覆人类以往生命观念的生物学革命。第一次在1967年的盛夏,由林恩·马古利斯(Lynn Margulis)挑起。马古利斯提出,复杂细胞不是经由“标准”的自然选择演化而来,而是通过一场合作共生的盛宴;细胞之间进行非常密切的合作,直到永久入住彼此体内。共生是指两个或更多物种之间一种长期的互动关系,通常伴随着物质和服务的交换。对微生物来说,物质交换涉及的是新陈代谢所需的、构成细胞生命的基础物质。马古利斯使用的术语是内共生(endosymbiosis):同样是交换物质,但这种合作方式极为亲密,合作一方干脆住到另一方的身体里面去了。马古利斯这些观点可以溯源到20世纪初期的研究,而且让人想起大陆漂移学说:非洲和南美洲的海岸线大致吻合,在地图上看,二者就像是曾经连在一起,后来才分开的。这种“幼稚”的想法,曾经被长期嘲笑为荒诞不经之谈。与之类似,复杂细胞的某些内部结构看起来很像细菌,而且好像是独立生长与分裂的。也许,解释就是那么简单——它们本来就是细菌!
与大陆漂移学说一样,这些思想都走在了时代的前面,直到分子生物学于20世纪60年代兴起,才给出了有力的论证。马古利斯以细胞内的两种专门构造为研究对象:线粒体和叶绿体。线粒体是细胞进行有氧呼吸的场所,食物在线粒体内氧化,并提供维持生命所需的能量。叶绿体是植物细胞进行光合作用的发动机,太阳能在这个场所中转化为化学能。这两种细胞器都保留了自身的微型基因组,其中只有几十个基因,编码几十种呼吸作用或光合作用所需的蛋白质。对这些基因测序后,真相终于明朗:简而言之,线粒体和叶绿体确实源自细菌。但它们不再是细菌,没有真正的独立性,因为绝大多数对它们自身存在必需的基因(至少有1500个)都储存在细胞核内。细胞核,才是细胞的“基因控制中心”。
马古利斯对线粒体和叶绿体的认识是正确的。到了20世纪80年代,反对者已经寥寥无几。但是她的野心远不止于此:在她看来,整个复杂细胞(现在的正式名称是真核细胞)都由共生细菌拼接而成。很多其他的细胞组成部分,尤其是纤毛(即列文虎克描述的“小脚”),同样起源于细菌(在马古利斯看来,纤毛起源于螺旋体)。她认为,一连串细菌共生合并造就了真核细胞。该理论后来被她命名为“系列内共生理论”。她认为还不只是单个的细胞,整个生物界的本质就是一张巨大的细菌协作网络:盖亚(Gaia)。她和詹姆斯·洛夫洛克(James Lovelock),成了“盖亚理论”的先驱。近些年,盖亚理论改头换面为“地球系统科学”(但摒弃了洛夫洛克原来的目的论成分),迎来了一次复兴;但是,马古利斯的观点,即认为真核细胞是细菌的集合体,从来没有多少实证基础。绝大多数真核细胞的内部构造,从形态上看并不像是起源于细菌,基因测序后更没有获得什么证据。所以,马古利斯在某些问题上是正确的,但在其他问题上几乎肯定错了。2011年,马古利斯因中风而过早去世。但她的斗士精神,她强烈的女性意识,她对达尔文主义式的竞争的排斥和她对阴谋论的轻信,都让她留下的智识遗产珠砾混杂。有人把她看作女权主义的英雄,有人认为她是我行我素的偏执狂。可悲之处在于,无论褒贬,这些争议大都远离科学。
第二次革命是种系发生学革命。这是一门关于基因族谱的学问,早在1958年,弗朗西斯·克里克(Francis Crick)就预见了这场革命的开端。克里克的预言,字里行间都带着他特有的从容自信:“生物学家应该意识到,很快我们就会迎来一门全新的学科,名字可能是‘蛋白分类学’,即对生物蛋白质氨基酸序列的研究,以及物种间的比较。这些序列是某种生物表型(phenotype)最细致的信息表达,它们之中可能隐藏着海量的生物演化信息。”他的预言果然成真。现代生物学的研究,很大程度上重心在于解开蛋白质和基因序列中隐藏的信息。我们已经不再直接比较氨基酸序列,而是比较DNA的字母序列(由它们编码蛋白质),因为这样准确度和灵敏度更高。尽管克里克有如此敏锐的预见性,但是他或者其他任何人,当时也完全想象不到,基因序列研究会牵出怎样的生命秘密。
卡尔·乌斯(Carl Woese)是一位历经坎坷的革命者。20世纪60年代,他低调开启了自己的研究工作,十年后才收获成果。当时,乌斯需要选择一个单一的基因来比较不同的物种。很明显,这个基因必须存在于所有物种之中,必须具有同一种功能。对细胞而言,这种功能必须非常基本、极为重要,任何细微改变都会遭到自然选择的惩罚。如果绝大多数变异都会被自然选择淘汰,那么留存下来的必然是基本不变或者说极端缓慢的演化,在漫长的岁月中改变非常小。如果我们要比较不同物种几十亿年间累积下的差异,就必须遵守以上选择标准。如果奏效,我们就能得到一张宏大的生命树图,一直追溯到生命起源。乌斯的计划就是这样志存高远。为了选出满足这些苛刻要求的基因,他开始研究细胞的基本功能,即细胞制造蛋白质的能力。
所有细胞都有核糖体。这是一种精巧的纳米机器,蛋白质的组装就是在核糖体上完成的。除了已经成为时代标志的DNA双螺旋,在信息时代的生物学领域,没有什么东西比核糖体更有象征意义了。它的构造还体现了一种人类思维难以测度的矛盾现象:比例。核糖体小到难以想象。细胞已经小到必须用显微镜观察;人类历史上的绝大多数时候,我们对细胞的存在都毫无概念。但核糖体比细胞还要小几个数量级,人类的一个肝细胞中就有1300万个核糖体。然而在原子维度上看核糖体,它又是非常巨大、极其复杂的超级建筑。由几十个基本部件组成,这些活动零件的运行精度,远超现代的自动化工厂流水线。这毫不夸张:核糖体首先与编码蛋白质的“穿孔带”代码脚本(即信使RNA)结合,然后按照序列逐个字母精确转译成蛋白质。核糖体捕获细胞质中的游离氨基酸作为基本构件,并把它们连成一条长链,顺序由代码脚本决定。核糖体的错误率大约为每10 000个字母出现一次错误,比人类高端制造业的废品率低得多。它们的工作效率大约为每秒钟处理10个氨基酸;由几百个氨基酸组成的蛋白质,一分钟内就能合成完。乌斯最终选择了核糖体的一个亚基,可以说是选了这台精密机器上的一个小零件,把它的编码基因作为比较对象。他比较了从细菌(例如大肠杆菌)到酵母再到人类等不同物种中的这一基因序列。
他的发现颠覆了我们的世界观。细菌和复杂真核生物之间的区别很明显,在表示亲缘关系的分支树图上,二者分属不同的分支大类。其中唯一的意外在于真核生物分支内部,即植物、动物和真菌之间的区别极小,但是,大多数生物学家仍在这个大类中投入了毕生的研究精力。真正出乎所有人意料的是,生命居然存在第三个域(最顶端的生物分类)。几个世纪以前,我们就已经知道这些简单细胞的存在,但一直把它们误认为细菌。它们的外观和细菌一模一样:同样微小,同样缺乏可以辨认的内部结构。但二者之间核糖体的区别,如同柴郡猫的神秘微笑,揭示了另有一种不同于细菌的生命存在—尽管同样缺乏复杂度。这种新的生物类群和细菌一样,没有真核生物的复杂形态,但它们的基因和蛋白质与细菌截然不同。这类生物被命名为古菌(archaea),因为当时的研究人员猜测,它们比细菌更古老。他们很可能猜错了,现代的研究认为,二者一样古老。然而在基因和生物化学层面,细菌和古菌之间的鸿沟就像细菌和真核生物(人类)之间一样巨大。在乌斯著名的“三域”生命树上,古菌域和真核生物域是“姊妹分支”,有较为接近的共同祖先。
古菌与真核生物在某些方面确实有很多相似之处,尤其是生物信息的流动(即读取基因序列并转化为蛋白质的方式)。古菌有几种精巧的分子机器,类似于真核生物的对等特征,只是少一些部件——也许这正是演化出真核生物复杂性的源头。乌斯不顾细菌和真核生物在形态上的巨大差异,而是把所有生物分成了地位相当的三个域,每个域内都经历了充分的演化,没有哪一支比其他任何一支更本源。他力主抛弃以前的术语“原核生物”(英文术语的原意为“出现细胞核之前”,适用于描述细菌和古菌),他的生命树图上也没有任何基因特征来反映“原核生物”这一划分。三个分支域都直接连回生命原点,拥有同一个神秘的祖先,不知如何就凭空出现了三支后代。乌斯晚年对生命最早期的演化持一种几乎是神秘主义的态度,呼吁用整体论来研究生命。这很讽刺,因为他本人掀起的生物学革命,恰恰是基于纯粹的还原论方法,即仅仅分析一个基因。细菌、古菌和真核生物确实是不同的类群,这点毫无疑问;乌斯的革命性开创也确实意义重大。但是他倡导的整体论方法,即以生物的整个基因组为研究对象,正在引发第三次细胞学革命。而这场革命,又推翻了乌斯自己的认识。
我们正处在第三次革命的进程之中。它的推理方法比较曲折,但带来的冲击最大。它的理论起源于前两次革命,特别致力于在二者之间建立联系。乌斯的生命树图,描绘了一个底层基因在三个生物域中的趋异演化关系。马古利斯描述的图景,则是不同物种的基因通过融合、捕获和共生行为,最终汇聚在一起的进程。如果把后者的想法也画成树图,会是一张汇聚图,而不是分支图,与乌斯的树图恰好相反。他们两个不可能全都正确!不过也不会全错。真相就隐藏在二者之间的某个位置,这是科学研究中常常遇到的情况。但不要以为这只是对两个理论的折中。目前正在成形的答案,比之前的两种结论都更加激动人心。
我们知道,线粒体和叶绿体确实源于细菌,它们是通过内共生作用融入细胞的;而真核细胞的其他部分,很可能通过常规方式演化而来。关键问题在于,融合究竟是什么时候发生的?叶绿体只存在于藻类和植物中,所以很可能是由这两类的某个共同祖先单独获得的。因此,这应该是一次较晚发生的事件。而线粒体存在于所有真核细胞之中(有一个实际上不是例外的“例外”,我们将在第一章中讨论),所以获得线粒体应该是一次较早发生的事件。有多早?或许我们可以换个问法:什么样的细胞获取了线粒体?以下是标准的教科书观点:它是一种十分复杂的细胞,类似于阿米巴原虫,是可以自由运动的捕食者,能够变形,并通过吞噬作用吞噬其他细胞。也就是说,与货真价实、构造完备的真核细胞相比,当初获得线粒体的细胞并没有多大差别。然而我们现在知道,这种看法是错误的。过去几年间,研究人员选择了更具代表性的物种,比较了大量基因,由此得出了毫不含糊的结论:那个宿主细胞是一个古菌,属于古菌域。所有的古菌都是原核生物,顾名思义,它们没有细胞核,没有性别,没有包括吞噬作用在内的一切复杂生命特征。这个宿主细胞在形态方面几乎没有任何复杂度可言。然而,不知如何,它捕获了后来变成线粒体的细菌。自那之后,它才演化出了所有的复杂特征。如果事实的确如此,那么复杂生命的单一起源很可能有赖于对线粒体的获取,是线粒体触发了复杂生命的狂飙演化。
复杂生命的起源,是发生在古菌宿主细胞和后来变成线粒体的细菌之间的内共生事件,而且只发生过一次。这个激进的理论,其实早在1998年就由演化生物学家比尔·马丁(Bill Martin)提出。马丁才华横溢,直觉敏锐,思维开阔。他发现了真核细胞不同寻常的基因嵌合现象,并据此提出了大胆的新理论。以发酵作用这条生物化学路径为例:古菌使用一种特定的方式,细菌则采用另一种,两种方式涉及的基因也大不一样。而真核生物会使用几种发现于细菌中的基因、几种发现于古菌中的基因,并把它们编织成一条精密的复合路径。这种错综复杂的基因嵌合现象,不止发酵作用有,复杂细胞中的所有生物化学过程几乎都是如此。从演化遗传学的角度来看,这种情况简直是“岂有此理”!
马丁考虑得十分周详。为什么宿主细胞从它的内共生体那里获取了这么多基因,并把它们深深地融入自己的基础架构之中,取代了很多自身原有的基因?马丁和米克洛什·米勒(Miklós Müller)共同提出的答案是“氢气假说”。他们认为,宿主细胞是一个古菌,能利用两种简单的气体生活:氢气和二氧化碳。内共生体(未来的线粒体)是一个有着多种代谢方式的细菌(这对细菌而言很普遍),为宿主细胞提供氢作为养料。马丁和米勒通过逻辑推理,一步步明确了这种共生关系的细节,解释了为什么一种原本依靠简单气体存活的细胞,后来会为了供养它的内共生体而变成以有机物为食。但这些都不是这里的关键。关键在于,马丁推测复杂生命起源于两种细胞之间的单一内共生事件。他认为,宿主细胞是古菌,不具有真核细胞的那些复杂特征;他还认为,从来就没有所谓的“中间型”,即不存在尚未获得线粒体的简单真核细胞;线粒体的获得和复杂生命的起源,本来就是同一事件。他还认为,真核细胞那些精巧繁复的特征,诸如细胞核、性和吞噬作用,全都发生在获取线粒体之后,是在这种独特的内共生状态下演化而来的。马丁的研究代表了演化生物学中最深刻的洞察力,本应该更加广为人知。但是它太容易被人与系列内共生理论混为一谈(后面我们会看到,内共生理论并没有提出马丁的这些推论),所以当时未能脱颖而出。然而过去20年间,马丁这些详尽的理论预言全都得到了基因组研究的证实。这真是一座生物化学逻辑严密推理的丰碑!如果诺贝尔奖单独设立生物学奖项,没有人比马丁更配得奖。
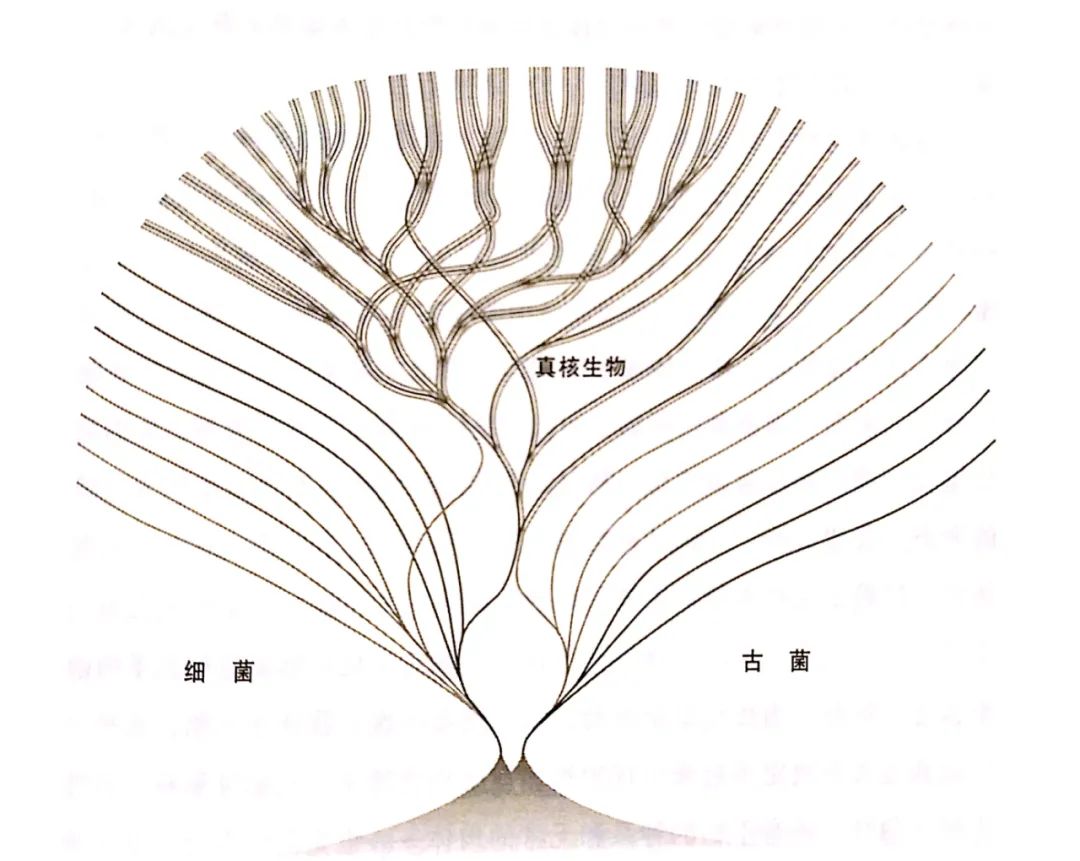
图1 显示复杂细胞嵌合起源的生命树(这是马丁于1998年绘制的复合生命树,依据的是对整个基因组的比较。细菌、古菌和真核生物三个域的关系如图所示。真核生物的起源是嵌合式的,古菌宿主和细菌内共生体的基因混合,宿主最终演化成了形态复杂的真核细胞,内共生体最终演化成了线粒体。某一类真核生物后来又获取了第二种细菌内共生体,最终演化成了藻类和植物的叶绿体。)
所以,我们现在又回到了起点。我们掌握了很多知识,但还是不知道为什么生命会是这样。我们知道复杂细胞起源于40亿年演化史中的单一事件:一个古菌和一个细菌的内共生(图1)。我们知道复杂生命的特征在这次结合之后才演化出现。但我们仍然不知道,为什么这些特征会出现在真核细胞中,在细菌和古菌中却没有留下演化的痕迹。我们不知道是什么力量限制了细菌和古菌:为什么它们的生物化学机制如此多姿多彩、基因多态性如此丰富、依靠气体和石头都能顽强生存,但一直保持着简单的形态?我们真正拥有的是一个新颖大胆的理论构架,可以依此继续探索。
我相信,线索就藏在细胞的生物能量生产机制中。这种怪异的机制从各方面限制了细胞,但很少有人理解这一点。本质上,所有的活细胞都通过质子(带正电荷的氢原子)回流来为自身提供能量,就像是某种电流——只是用质子代替了电子。我们通过呼吸作用氧化食物而获得的能量,被用来把质子泵过一层膜,在膜的另一边形成质子蓄积。从这个“水库”回流的质子可以为细胞工作供能,如同水电站的涡轮电机。这种利用跨膜质子梯度为细胞供能的奇特机制,发现之初完全出人意料。彼得·米切尔(Peter Mitchell)是20世纪最具独创性的科学家之一,他于1961年首先提出了这一理论,并在此后的30年间逐渐将其完善。米切尔的学说被认为是自达尔文以来最“反直觉”的生物学理论,可与爱因斯坦、海森堡和薛定谔的物理学思想相媲美。现在,我们对质子动力的工作方式理解得十分详尽,已经深入到蛋白质层面。我们还发现,所有的地球生命,无一例外都利用质子梯度供能。质子动力是生命不可或缺的成分,就像通用遗传密码。然而,这种反直觉的能量利用机制最初是怎么演化出来的,我们几乎一无所知。在我看来,这就是位于当代生物学核心的两大未知问题:为什么生命以如此令人困惑的路径演化?为什么细胞的供能方式如此古怪?
写作本书就是为了尝试回答以上两个问题,而且我相信二者紧密相关。我希望说服读者,演化是围绕能量进行的,我们必须考虑能量才能理解生命的各种特征。我希望向读者展示,能量与生命从一开始就密不可分,地球生命的基本特征源于一颗躁动行星的能量失衡。生命的起源由能量流推动,质子梯度对细胞的出现至关重要,但是对质子梯度的利用又限制了细菌和古菌的结构。这些限制条件主宰了细胞之后的演化历程,细菌和古菌虽然在生物化学方面花样百出,却一直保持着简单的形态。我想证明,一次罕见的内共生事件,即一个细菌入住一个古菌体内,打破了这些限制,使复杂细胞的演化成为可能。一个细胞在另一个活细胞内生活并逐渐融合,这是很难实现的变化;形成这种关系的困难程度,解释了为什么复杂生命的起源只有一次。我还想证明,这种密切的共生关系决定了后来出现的很多复杂细胞特征,包括细胞核、有性生殖、两性,还有不朽的种系和无常的肉体—也是有限寿命和基因预定死亡的源头。最后,从能量角度思考生命能让我们认识人类自身的生物学特性,特别是演化过程中深层次的取舍权衡:生殖力和年轻时的健康,代价是衰老和疾病。我认为,这些见解能够帮助人类增进健康,至少能加深对健康的理解。
本文节选自《复杂生命的起源》,贵州大学出版社2020年10月版。
作者简介:
尼克·莱恩(Nick Lane),演化生物学家,英国伦敦大学学院教授。他的研究方向为演化生物学与生物能量学,聚焦于生命的起源与复杂细胞的演化。他还是伦敦大学学院线粒体研究学会的创始成员,也是生命起源研究计划的项目领头人。2010年,他以《生命的跃升》荣获英国皇家学会科学图书奖。尼克本人因为在分子生物学研究上的卓越贡献,于2015年荣获英国生物化学学会奖。他不仅深耕自己的学术研究领域,还孜孜不倦地参与公众科学普及。2016年,尼克因为在科学传播上的深入付出而荣获英国皇家学会迈克尔·法拉第奖章。
版权说明:欢迎个人转发,任何形式的媒体或机构未经授权,不得转载和摘编。转载授权请在「返朴」微信公众号内联系后台。
原标题:《生物演化史上的“黑洞”,可能需要物理学来填补丨展卷》
本文为澎湃号作者或机构在澎湃新闻上传并发布,仅代表该作者或机构观点,不代表澎湃新闻的观点或立场,澎湃新闻仅提供信息发布平台。申请澎湃号请用电脑访问http://renzheng.thepaper.cn。



- 报料热线: 021-962866
- 报料邮箱: news@thepaper.cn
互联网新闻信息服务许可证:31120170006
增值电信业务经营许可证:沪B2-2017116
© 2014-2024 上海东方报业有限公司

